İZMİR İLİNDEKİ FARKLI KAYNAKLARDAN İZOLE EDİLEN BAZI MAVİ-YEŞİL ALGLERİN (SİYANOBAKTERİLERİN) MOLEKÜLER ÖZELLİKLERİNİN VE TOKSİNLERİNİN ARAŞTIRILMASI
|
|
|
- Dilara Altun
- 5 yıl önce
- İzleme sayısı:
Transkript
1 EGE ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ (DOKTORA TEZİ) İZMİR İLİNDEKİ FARKLI KAYNAKLARDAN İZOLE EDİLEN BAZI MAVİ-YEŞİL ALGLERİN (SİYANOBAKTERİLERİN) MOLEKÜLER ÖZELLİKLERİNİN VE TOKSİNLERİNİN ARAŞTIRILMASI Zeliha DEMİREL Tez Danışmanı : Prof. Dr. Atakan SUKATAR Biyoloji Anabilim Dalı Bilim Dalı Kodu : Sunuş Tarihi : Bornova-İZMİR 2010
2 ii
3 iii Zeliha DEMİREL tarafından Doktora tezi olarak sunulan İzmir ilindeki farklı kaynaklardan izole edilen bazı mavi-yeşil alglerin (siyanobakterilerin) moleküler özelliklerinin ve toksinlerinin araştırılması başlıklı bu çalışma E.Ü. Lisansüstü Eğitim ve Öğretim Yönetmeliği ile E.Ü. Fen Bilimleri Enstitüsü Eğitim ve Öğretim Yönergesi nin ilgili hükümleri uyarınca tarafımızdan değerlendirilerek savunmaya değer bulunmuş ve tarihinde yapılan tez savunma sınavında aday oybirliği/oyçokluğu ile başarılı bulunmuştur. Jüri Üyeleri: İmza Jüri Başkanı : Prof. Dr. Atakan SUKATAR... Raportör Üye : Prof. Dr. İsmail KARABOZ... Üye : Prof. Dr. Meltem CONK DALAY... Üye : Prof. Dr. Arif GÖNÜLOL... Üye : Doç. Dr. Güven ÖZDEMİR...
4 iv
5 v ÖZET İZMİR İLİNDEKİ FARKLI KAYNAKLARDAN İZOLE EDİLEN BAZI MAVİ-YEŞİL ALGLERİN (SİYANOBAKTERİLERİN) MOLEKÜLER ÖZELLİKLERİNİN VE TOKSİNLERİNİN ARAŞTIRILMASI DEMİREL, Zeliha Doktora Tezi, Biyoloji Bölümü Tez Yöneticisi: Prof. Dr. Atakan SUKATAR Eylül 2010, 151 sayfa Toksik mavi-yeşil alglerin (siyanobakteriler) nehirler, göller ve denizsel çevrelerde bulunduğu rapor edilmektedir. Fakat toksin üreten mavi-yeşil algler çoğunlukla sıcak su kaynaklarda araştırılmamıştır. Bu nedenle bu çalışmada İzmir ilindeki 5 farklı (Balçova, Zeytindalı, Karakoç, Şifne ve Gülbahçe) sıcak su kaynaklarındaki toksik siyanobakteriler ve bunların siyanotoksinlerinin varlığı araştırılmıştır. 10 ipliksi siyanobakteri türü 16S rdna dizisi çıkarılmış ve bu dizilerin filogenetik ağacı ve GenBank yapıları oluşturulmuştur. İzmir ilindeki termal alanlardaki siyanobakteri kültürlerinden 10 (Spirulina subsalsa, Geitlerinema, Leptolyngbya, Lyngbya, Oscillatoria, Pseudoscillatoria, Phormidium sp.) tür izole edilmiştir. İzolatların içeriğinin toksik aktivitesi mikrosistin enzim bağlı immunosorbent deneyi (ELİSA) ve polimeraz zincir reaksiyonun (PCR) da spesifik hedef pirmerlerin mcya geninin çoğaltılması kullanılarak doğrulanmıştır. ELİSA sonucunda siyanobakterilerin mikrosistin içeriğinin konsantrasyonu 0,16 ppb belirlenen limitten az olduğu ortaya çıkarılmıştır. Mikrosistin sentez kompleksindeki mcya gen bölgesi 5 örnekte belirlenmiştir. Bu çalışma ile ilk kez İzmir ilindeki termofilik siyanobakterilerin mikrosistin bulundurduğu rapor edilmiştir. Anahtar sözcükler: Termofilik siyanobakteri (mavi-yeşil alg), DNA izolasyonu, PCR, ELİSA, toksin
6 vi
7 vii ABSTRACT INVESTIGATION OF BLUE-GREEN ALGAE(CYANOBACTERIA) MOLECULER PROPERTIES AND TOXIN ISOLATED FROM IZMIR DIFFERENT SPRINGS DEMIREL, Zeliha PhD in Department of Biology Supervisor: Prof. Dr. Atakan SUKATAR September 2010, 151 pages Toxic blue-green algae (cyanobacteria) are well reported in rivers, lakes and marine environments, but the toxin production of blue-green algae in hot springs is largely unexplored. Therefore, the present study investigated the presence of toxic cyanobacteria and cyanotoxins in 5 different hot springs (Balcova, Zeytindal, Karakoc, Sifne and Gulbahce) in Izmir. 16S rdna sequences from 10 filamentous cyanobacteria were obtained and phylogenetic tree from these sequences and sequences available in GenBank was constructed. Ten cyanobacterial cultures isolated from thermal places in Izmir, Spirulina subsalsa, Geitlerinema, Leptolyngbya, Lyngbya, Oscillatoria, Pseudoscillatoria, Phormidium sp. contained with toxic activity confirmed using microcystin enzyme-linked immunosorbent assay (ELISA) and polymerase chain reaction (PCR) with specific primers targeted to the condensation domain of mcya gene. The results of an ELISA revealed that cyanobacteria contained microcystin at concentrations below detection limits of 0,16 ppb. The gene mcya from the microcystin synthesis complex was detected in five samples. This is the first report of the presence of microcystin thermophilic cyanobacteria in Izmir. Keywords: Thermophilic cyanobacteria (Blue-green algae), DNA isolation, PCR, ELISA, toxin
8 viii
9 ix TEŞEKKÜR Bu çalışmada manevi desteğini esirgemeyen sayın hocam Prof. Dr. Atakan SUKATAR a ve maddi olarak destekleyen Türkiye Bilimsel ve Teknik Araştırma Kurumu TÜBİTAK a ve Ege Üniversitesi Bilim-Teknoloji Uygulama ve Araştırma Merkezi EBİLTEM e teşekkür ederim. Araştırmamızda gerekli cihazların kullanımına izin veren hocalarım Doç. Dr. Güven ÖZDEMİR, Doç. Dr. İhsan YAŞA, Doç. Dr. Remziye DEVECİ ve Prof. Dr. Figen ZİHNİOĞLU na destekleri için teşekkür ederim. Tezin biçimlenmesinde değerli yardımlarından dolayı Araş. Gör. Dr. Ali KOÇYİĞİT, Araş. Gör. Dr. Savaş İZZETOĞLU, Araş. Gör. Ali ZEYTÜNLÜOĞLU ve Uzman Biyolog Ebru TEKİN e teşekkürü bir borç bilirim. Beni destekleyen ve her zaman sabırla yanımda olan biricik aileme ve arkadaşlarıma çok teşekkür ederim.
10 x
11 xi Ç NDEK LER Sayfa ÖZET... v ABSTRACT...vii TE EKKÜR...ix EK LLER D Z N...xv Ç ZELGELER D Z N...xix S MGELER VE KISALTMALAR D Z N...xxi 1.G R ÖNCEK ÇALI MALAR Siyanobakteriyel Toksinler Mikrosistin Nodularin Anatoksinler Saksitoksinler (Paralitik Kabuklu Zehirler (PSPs)) Cylindrospermopsin Lipopolisakkarid (LPS) endotoksinler MATERYAL VE YÖNTEM...22
12 xii Ç NDEK LER (devam) Sayfa 3.1 Organizma Kültür ortam Stok kültür haz rlanmas Fiziksel ko ullar Termal alandaki su özellikler Mikroskop Kimyasallar Moleküler Tan lanmas Polimeraz Zincir Reaksiyonu Sekans analizi Denature Edici Gradient Jel Elektroforez Enzim Ba l mmuno Sorbent Analizi (EL SA) SONUÇLAR VE TARTI MA ÖNER LER KAYNAKLAR
13 xiii Ç NDEK LER (devam) Sayfa ÖZGEÇM...151
14 xiv
15 xv EK LLER D Z N ekil Sayfa 2.1 Mikrosistin ve nodularin izoformlar Mikrosistin ve nodularin gen yap lar Nodularia spumigena nodularin gene yap s Anatoksinlerin genel yap lar Oscillatoria PCC 6506 anatoksin-a ve homoanatoksin-a biyosentezinden sorumlu olan tahmini gen kümelerinin organizasyonu Saksitoksinlerin genel yap lar Paralitik Kabuklu Zehirler biyosentez yap s (sxt) Cylindrospermopsinin genel yap s C. raciborskii T3 sxt gen kümelerin yap sal organizasyonu Stok kültür haz rlanmas ve türlerin inkübasyon ortamlar ifne Termal Otel (Havuz) Balçova Termal Otel (S cak su ç k ) Zeytindal Termal Otel (S cak su ç k ) Karakoç Termal Havuzu ve s cak su ç k...28
16 xvi EK LLER D Z N (devam) Sayfa 3.6 Tarihi Urla-Gülbahçe Termal Havuzu Agaroz jel elektroforezi DGGE (Scie-Plas) 60 ºC de 80 V yürütülmesi Urla 1 (U sal) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning Elektron mikroskobu Karakoç 1 (KY) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu Karakoç 2 (KK) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu Balçova 1 (Bal ip) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu ifne ( if) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu Balçova 2 (Bal pse) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu Zeytindal (ZD) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu Urla 2 (U ip) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu Urla 3 (U pse) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu... 52
17 xvii EK LLER D Z N (devam) Sayfa 4.10 Urla 4 (U osc) foto raflar a. I k mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu Agaroz jel üzerinde kitlerden elde edilen DNA lar n kar la t r lmas ZRFungal/Bacterial DNA Kiti ile DNA izolasyonu Nanohelix kiti kullan larak 16SF-23SR bölgesinin ço alt lan görüntüsü Crimson kiti kullan larak 16SF-23SR bölgesinin ço alt lan görüntüsü Nanohelix hypersense kiti kullan larak 27F-809R bölgesinin ço alt lan görüntüsü Nanohelix hypersense kiti kullan larak 16SF-23SR bölgesinin ço alt lan görüntüsü Nanohelix hypersense PcβF-PCαR bölgesinin ço alt lan görüntüsü Nanohelix hypersense 27F-1492R bölgesinin ço alt lan görüntüsü DGGE de if, U pse, KK, KY türlerinin kar la t r lmas DGGE de KK, ZD, if türlerinin kar la t r lmas DGGE de KK, U ip, ZD türlerinin kar la t r lmas DGGE de KY, KK, Bal pse, Bal ip türlerinin kar la t r lmas DGGE de U pse, U osc, U ip, U sal türlerinin kar la t r lmas...133
18 xviii EK LLER D Z N (devam) Sayfa 4.24 DGGE de Urla, ifne, Zeytindal ve Karakoç termallerinin izole edilmeyen kar k kültürlerinin kar la t r lmas EL SA platenin A. Durdurma çözeltisi eklenmeden önce B. Durdurma çözeltisi eklendikten sonraki görüntüsü Saksitoksin gen bölgelerinin görüntüsü Mikrosistin A gen bölgelerinin görüntüsü Nodularin gen bölgesinin görüntüsü
19 xix Ç ZELGELER D Z N Çizelge Sayfa 2.1 Siyanobakteriyel toksin gruplar ve kaynaklar zole edilen özgün türler ve kodlar Blue Green Algae (BG-11) ortam kompozisyonu ifne Termal Suyuna ait kimyasal kompozisyon Balçova Termal Suyuna ait kimyasal kompozisyon Zeytindal Termal Suyuna ait kimyasal kompozisyon Lineer DNA n n ayr lmas için önerilen jel yüzdesi Bakteri ve siyanobakteri belirlenmesi için seçilen primer çiftleri DGGE de kullan lan siyanobakteri primer çiftleri Siyanobakteri toksinleri için seçilen primer çiftleri DGGE Jel kompozisyonu F-809R için dizi analizi sonuçlar F-1492R Dizi analizi sonuçlar SF-23SR Dizi analizi sonuçlar Pc βf-αr dizi analizi sonuçlar F-R dizi analizi sonuçlar...122
20 xx Ç ZELGELER D Z N (devam) Sayfa 4.6 EL SA sonuçlar
21 xxi S MGELER VE KISALTMALAR D Z N Simgeler Aç klama kb Kilobaz. M Marker. Mcy Mikrosistin gen bölgesi. K saltmalar PCR Polimeraz zincir reksiyonu. DGGE Denature gradient jel elektroforezi. EL SA Enzim ba l immunosorbent analizi. NRPS Non-ribozomal peptid sentetaz. PKS Poliketit sentetaz.
22 xxii
23 1 1. GİRİŞ Dünya çevresindeki siyanobakterilerin (mavi-yeşil alglerin) çoğu tatlı su ve denizlerde yayılış göstermektedir. Siyanobakterilerin tanımlanmasındaki geleneksel yöntem sadece mikroskop kullanılmasıyla morfolojik tayin olmasından dolayı toksinler direkt analizlerle birleştirilmiştir. Morfolojik tanımlamalarda toksik ve toksik olmayan siyobakterilerin ayrımı zor olmaktadır. Bazı genuslar hem toksik hem de toksik olmayan üyelere sahiptir. DNA ya dayanan belirleme yöntemlerin potansiyel özgünlüğü (hedef genin içerdiği toksin biyosentezi), duyarlılığı ve hızlı olması popüler hale gelmesine neden olmuştur. Son yıllarda, özellikle yurt dışında, sistematik çalışmalarda kullanılan morfolojik tayinlerin yetersiz olduğu durumlarda moleküler düzeyde çalışmalar yapıldığı görülmektedir. Bu çalışmada, ülkemizde termal ortamlarda gelişim gösteren siyanobakteriler ile ilgili moleküler düzeyde yeterli çalışma bulunmaması nedeniyle termal alanlardaki çeşitliliğin belirlenmesi hedeflenmiştir. Elde edilen türlerin toksik ve toksik olmayanlarının moleküler analizlerle desteklenmesi amacıyla gerçekleştirilecek olan bu çalışmanın bu alandaki boşluğu dolduracağını düşünmekteyiz. Yurtdışında çok fazla yayın ve çalışma bulunmasına karşın ülkemizde güzellik merkezlerinde ve kaplıcalarda kullanılan sularda ve çamurlarda gelişen siyanobakteri türlerinin belirlenmesi, bunların toksinlerinin bulunup bulunmadığı ve varsa toksinlerinin miktarlarının tespiti ülkemizde henüz araştırılmamıştır. Bu bilgilerin ışığında ülkemizde bu alandaki eksikliğin tamamlanacağını ve diğer çalışmalara kaynak olarak kullanılabileceği düşüncesindeyiz. Bu amaçla; ülkemiz sularındaki termofilik siyanobakteri türlerini, toksinlerini ve toksin miktarlarının belirlenmesi yönünde çalışma yapılmıştır. İzmir ili ve çevresindeki sıcak su kaynaklarında gelişen siyanobakteriler izole edilerek saflaştırılmış ve üretilen örneklerin DNA sının polimeraz zincir reaksiyonu (PCR) ile belirlenmesinden sonra, dizi analizine gönderilerek türler
24 2 tanımlanmıştır. Bu sayede morfolojik ve moleküler tanılamaları karşılaştırmalı olarak yapılmıştır. Siyanobakterilerin toksik ve toksik olmayan türleri bulunduğu için, tanımlanan türlerin toksin içerip içermediği ve miktarları Mikrosistin Enzim Bağlı İmmunosorbent Analizi (ELİSA) test kiti kullanılarak belirlenmiştir. Toksin içeren türlerin DNA sı toksin gen bölgesini (mcy) kapsayan primerlerden yararlanarak PCR ile çoğaltılmış ve elektroforezde görüntülenmiştir.
25 3 2. ÖNCEKİ ÇALIŞMALAR Günümüze kadar Türkiye sularında sistematik açıdan birçok araştırma yapılmış ve Ege Bölgesi termal sularının alg florasının çoğu ortaya çıkarılmıştır (Güner, 1966; Güner, 1967; Güner, 1970; Ulcay, 2005; Ulcay et al., 2007; Yüksel, 2009). Cadirci et al tarafından gerçekleştirilmiş olan bir çalışmada Balçova- İzmir den termofilik bir siyanobakterinin izolasyonu ve moleküler identifikasyonu yapılmıştır. Yüksel (2009) master tezinde İzmir ilindeki Balçova, Şifne ve Zeytindalı kaplıcalarından 4 farklı siyanobakteri türü izole etmiş ve bir türü moleküler olarak tanımlamıştır. Siyanobakteriler, 16S rrna ve klorofil-a içeren, karbondioksit ve atmosferik azotu fiske edebilen, fotosentez yapma yeteneği olan, organik karbon ve oksijen üretebilen tek prokaryot organizma grubudur. Mavi-yeşil algler olarak da adlandırılırlar ve çoğunlukla sucul veya karasal habitatlarda yaşarlar. Siyanobakteriler ekolojik, evrimsel ve ekonomik önemi olan fotosentetik organizmalardır (Carmichael, 1994; Mejean et al., 2010). Fotosentetik olmaları nedeni ile üretim sistemleri mikroalglerle aynıdır. Mikroalg ve siyanobakteriler fitoplanktonu oluşturur. Balıklar ile diğer sucul organizmalar için besin maddesi olarak kullanılmalarından dolayı doğadaki besin zincirinin başlangıç noktasını oluştururlar. Mikroalgal biyoteknolojiyi konu alan pek çok kaynakta siyanobakteriler, mikroalglerle birlikte değerlendirilir (Borowitzka and Borowitzka,1992; Sasson, 1997). Birçok siyanobakteri hücresi veya filamenti kendilerini bir arada tutan müsilajlı kılıflar üretirler. İpliksi siyanobakterilerde, filamentlerin fragmantasyonu genelde hormogonyumların oluşması ile meydana gelmektedir. Bu yapı filamentlerden koparak, kayma hareketi ile ayrılır. Bazı türlerde, sporlar ya da akinetler oluşur. Sporlar ve akinetler organizmayı, karanlık, kuruma ve donmaya karşı korur. Sporlar ve akinetler, kalınlaştırılmış dış hücre duvarına sahip hücrelerdir ve organizma bu dış duvarın tekrar yıkılması ve ardından yeni vejetatif filamentin dışarı çıkması ile yeniden gelişmeye başlar. Bununla birlikte bir çok
26 4 siyanobakteri rölatif olarak kurumaya ve düşük sıcaklıklara dayanıklıdır (Madigan et al., 2000). Siyanobakteriler klorofilin sadece tek formuna, klorofil-a ya sahiptirler. Ayrıca hepsinde karakteristik biliprotein pigmentleri olan, fotosentezde yakalayıcı pigment olarak fonksiyon gösteren fikobilinler mevcuttur. Fikobilinlerin bir sınıfı olan fikosiyaninler, mavi-yeşil renklidir ve klorofil-a ile birlikte siyanobakteriye mavi-yeşil rengini verirler. Bazı siyanobakteriler ise kırmızı renkli fikobilin olan fikoeritrini üretirler. Bu pigmente sahip olan türler ise kırmızı ya da kahve renklidir. Siyanobakterilerde görülen sitoplazmik yapılar arasında gaz vakuolleri de bulunur. Özellikle açık sularda yaşayan planktonik türlerde, yüzebilirliği ayarlama fonksiyonundan dolayı çok yaygındır. Bu sayede fotosentez için gerekli optimum ışığı alabilecekleri seviyede kalabilirler (Madigan et al., 2000). Fotosentez, fotosentetik bakteriler ve yüksek bitkiler gibi bütün fotoototrofik organizmalar tarafından gerçekleştirilen, ışık enerjisinin kimyasal enerjiye dönüştüğü metabolik bir işlemdir. Fotosentez olayı iki evreyi içerir. Işık reaksiyonları, ışık enerjisinin yakalanmasını ve ATP ye dönüşümünü gerektirir. Bu işlem, ışık aracılığı ile H 2 O dan NADP ye elektron taşınımı ile gerçekleşir. Elektron taşınımında reaksiyonların olduğu yer hücre içindeki tilakoid zarlardır. Yüksek bitkilerden farklı olarak, bu zarlar siyanobakterilerde organize bölgeler değillerdir. Bu reaksiyonlar yüksek bitkilerde kloroplast içinde gerçekleşirken siyanobakterilerde sitoplazma içinde oluşmaktadır. Tilakoidler klorofil a içerirlerken klorofil b ve c bulunmaz. Siyanobakterilerde rezerv polisakkarit, glikojene benzer siyanofisin nişastasıdır (Vonshak, 1997). Siyanobakteri hücre çeperi birkaç tabakadan oluşur. Siyanobakteri hücre çeperinin temel bileşeni peptidoglukan(=murein,glikopeptid=mukopeptid) dır. Sadece en içteki iki tabaka tüm siyanobakterilerde aynıdır. Dışta bulunan çeperin yapısı, çevre koşullarına ve salgılanan müsilaj miktarına göre değişim gösterir. Siyanobakterilerin hücre duvarının yapısında peptidoglikan mevcuttur, ve bu nedenle Gram-negatif bakterilere benzemektedirler (Madigan et al., 2000). Siyanobakteri hücre çeperi yapısının temel olarak gram negatif bakterilerin hücre çeperi ile büyük benzerlik göstermesi nedeniyle bu iki grup arasında bir
27 5 akrabalığın olduğu düşünülmektedir. Hücre çeperinin dışında bulunan müsilaj katman (örtü ya da kapsül) fibrilli yapıdadır. Bu katmanın görevi hücreleri kuraklığa karşı korumaktır. Ayrıca hücrenin kayarak ilerlemesinde de işlev görür. Kılıflar bazen renklidir. Kırmızı katmana sahip algler genellikle yüksek asidik topraklarda, mavi katmana sahip olan algler ise bazik topraklarda gelişim gösterirler. Sarı ve kahverengi örtüye sahip olan algler ise yüksek tuz konsantrasyonlu bölgelerde bulunurlar (Sukatar, 2004). Toksik algler (ökaryotik ve prokaryotik (mavi-yeşil algler)) hem tatlı sularda hem de deniz sistemlerinde bulunmaktadır. Bu organizmaların içinde yer alan siyanobakteriler ani çoğalma gösterdiklerinde (algal bloom), göl ve havuz sularının renklerini kırmızı ve yeşile dönüştürebilmektedir. Bu alglerin oluşturduğu toksinlere kısaca siyanotoksinler denilmektedir, siyanobakterilerin genellikle ürettiği toksinler sekonder metabolitlerdir ve sayıca çok fazla organizma tarafından üretildiği bilinmektedir. Ouellette ve Wilhelm 2003 tarafından yapılan çalışmada, moleküler yöntemler kullanılarak toksik siyanobakterilerin belirlenmesi hedeflenmiştir. Toksik siyanobakteriler; siyanobakteriyel ekoloji, siyanotoksin üretimi, toksikolojisi ve su yönetimi gibi konularla ilişkilendirilmektedir (Quellette and Wilhelm, 2003). Geleneksel olarak siyanobakteriler sadece mikroskobik olarak tanımlanmasının yanı sıra toksinlerin analizleri de tanımlanmaktadır. Morfolojik tanılamalar ile toksik ve toksik olmayan türlerin birbirinden ayrımı zor olmaktadır. Çünkü bazı cinsler hem toksik hem de toksik olmayan türleri içermektedirler. DNA tabanlı belirleme yöntemlerinin popüler olmasında, potansiyel özgünlük (hedeflenen genlerin toksin biyosentezinde yer alması), duyarlı ve hızlı sonuçlar vermesi gösterilmektedir (Quellette and Wilhelm, 2003). Siyanobakteriyel biyoçeşitliliğin değerlendirilmesinde moleküler yaklaşımlarda PCR dan bağımsız ve PCR tabanlı olmak üzere iki temel yöntem ileri sürülmüştür. PCR dan bağımsız yaklaşımlar; GC içeriği, DNA hibridizasyonu, DNA mikroarray gibi teknikler içerirken, PCR tabanlı yaklaşımlar; DNA klonlama ve dizileme, DGGE, TGGE (Termal gardientli jel elektroforezi), SSCP (Tek Zincir Konformasyon Polimorfizmi), RFLP
28 6 (Restriksiyon Parçası Uzunluk Polimorfizmi), ARDRA (amplified rdna restriksiyon analizi), T-RFLP (Terminal Restriksiyon Parçası Uzunluk Polimorfizmi), RISA (ribozomal intergenik spacer analizi), ARISA (otomatikleştiriliş ribozomal intergenik spacer analizi), RAPD (random amplifiye edilmiş polimorfik DNA). Siyanobakterilerin biyolojik çeşitliliğinin moleküler değerlendirilmesinde sıklıkla kullanılan belirleyiciler, 16S rdna, fikosiyanin lokusu, nif geni, rpo geni, ITS gen bölgesi olarak açıklanmıştır (Kumari et al., 2009). Tatlı sularda bulunan siyanobakterilerin ani çoğalmalarının dünya genelinde artmasıyla ilgili olarak birçok türün ürettiği toksinler insanlar ve diğer organizmalar için zararlıdır. Siyanobakterilerin 1500 den daha fazla türden en az 30 tanesinin toksik bileşikler ürettiği bilinmektedir. Potansiyel toksik türlerin arasında bazı kimyasallar ve toksikolojik farklı bileşikler izole edilip, saflaştırılmış ve karakterizasyonları yapılmıştır (Saker et al., 2005). Mikrobiyal kommünitelerin genetik özelliklerinin araştırılmasında en iyi tekniklerden biri 16S rrna PCR-DGGE (Polimeraz Zincir Reaksiyonu-Denature Gradient Jel Elektroforezi) yöntemidir. Bu yöntemde ayrılan küçük DNA fragmentlerinin (max. uzunluk 1000bp) erime özellikleri nedeniyle farklı diziler izlenmektedir. Genellikle ileri primerin 5 sonuna bir GC clamp eklenerek çift sarmallı fragmentlerin komple denaturasyonu engellenmektedir. İleri veya geri primer üzerindeki GC clambın pozisyonu kalıpların kalitesi ile ilgili olmasına rağmen daha önceden araştırılmamıştır (Boutte et al., 2006). DGGE den elde edilen bantların sayıları, pozisyon ve yoğunlukları doğal örneklerin çeşitliliklerinin belirlenmesinde kullanılmaktadır. Bazı önyargılarda DGGE kalıplarının güçlü açıklamalar oluşturamadığı rapor edilmiştir. Örneğin; heterodupleks formülasyonunda, amplikasyon hataları, 16S rrna operonlarındaki multipli kopyaların bulunması, DNA nın farklı fragmentlerindeki co-migration (eş göçler) ve DNA fragmenti içinde bazı erime alanlarının bulunmasında olduğu gibi. Ama yine de bu problemlerin bazıları çözülebilmesinde alınan bantların reamplikasyon ve dizilemenin izlenmesi, DNA fragmenti analizindeki polimorfizmin tanımlanan organizmalarda anlaşılmasıyla gerçekleşmektedir (Nübel et al., 1997).
29 7 Nübel et al., (1997) siyanobakteri primerleri olan CYA359F (ileri), CYA781R (a) ve CYA781(b) (geri) için spesifik amplikasyon 379 bp 16S rrna gen dizisini dizayn etmiştir. CYA781R (a) ve CYA781(b) nin farkı iki polimorfizimin temeldeki durumunda 7 ve 23 pozisyonlardır (5 den 3 ). Bu primerlerin avantajı verdiği PCR ürünlerinin V3 ve V4 bölgelerine benzemektedir. Bu yüzden önemli filogenetik bilgiler içermektedir. Aslında pimerler birçok DGGE çalışmalarında çevresel örneklerin siyanobakteriyel çeşitliliği araştırılmasında değiştirilmeden kullanılmıştır. Doğadaki mikrobiyal kommünitelerin anlaşılması için, mikroorganizmaların 16S rrna dizilerinin karşılaştırılma analizlerinin dünyada uygulamasının sebebi, bütün organizmalarda ribozomal RNA moleküllerinin bulunmasıdır ve bu organizmaların arasındaki ilişkilerinin anlaşılmasını sağlamaktadır. rrna molekülleri yüksek oranda korunan dizi kısımları ile değişken bölgelerinin karşılaştırılmasından oluşmaktadır. Genel olarak temel rrna kalıpları bütün filogenetik kalıplar boyunca korunmaktadır. Böylece dizilerin universal izleri tanımlanabilmektedir (Cho et al., 2003). Cho et al., (2003) yaptıklaarı çalışmada iki PCR tabanlı-16s rdna ile başlayarak ARDRA ve DGGE yardımıyla yer altı sularının bakteriyal kommünite yapılarının karakterizasyonunu çalışılmıştır. Kommünite çeşitliliğinin ölçülmesinde çok basit yöntem olan ARDRA kullanılmıştır. Bu yöntem temelde çevre örneklerinin popülasyonun rdna dizilerinin doğrudan arttırması için restriksiyon parçalaması yapılmasına rağmen bu yöntemin karmaşık kommüniteler içindeki spesifik filogenetik grupların tanımlanmasında ayırım gücü sınırlıdır. ARDRA restriksiyon kenarlarının dizi bilgisinde kullanılabilir. Son zamanlarda, PCR denature edici gardient jel elektroforezi (PCR-DGGE) kullanılmaktadır. Yeni bir yöntem olan PCR-DGGE yöntemi ile çevresel örneklerdeki bakteriyel populasyonların filogenetik çeşitliliği araştırılmaktadır. Bu yöntemde toplam mikrobiyal DNA yeraltı sularından ekstrakte edilmekte ve bakterilerin 16 srna genleri PCR da universal öbakteriyal primerler ile
30 8 çoğaltılmaktadır. Aynı uzunluktaki PCR ürünlerinin DGGE de erime özelliğiyle farklı iç dizileri ayrılması sağlanmaktadır. DGGE, DNA fragmentindeki aynı uzunluktaki diziler arasındaki farklılıkların belirlendiği elektroforetik bir yöntemdir. Bu fragmentler denature edici gardient jelde bunların farklı denatürasyon (melting-erime) görüntüsüyle ayrılabilmektedir. Teorik olarak bu ayrılma görüşü ilk olarak Fisher ve Lerman (1983) tarafından tanımlanmıştır. Akrilamid jelde denatürasyon şartları üre ve formamide ile sağlanmaktadır. Çözelti, % 100 kimyasal denatürasyonu sağlamak için 7 M üre ve % 40 formamid içermelidir. Düşük ve yüksek denatürasyon çözeltileri hazırlanır ve bu çözelti akrilamid ile karıştırılır ve jel kasketini içine gradient karıştırıcı yardımıyla dökülmelidir. Elektroforez sabit sıcaklıklarda 55ºC ve 65ºC arasındaki sıcaklıklarda genellikle 60ºC de yürütülmektedir (Ercolini, 2004). DGGE jelde çift iplikli DNA fragmentleri artan denatürasyon çevresine maruz kalmaktadır. Böylece DNA artan konsantrasyondaki denatürasyon ajanları ve kısmi erime ile farklı bölgelere kısaca eriyen kısımlarla çarpışmaktadır. Erime sıcaklığı (Tm) alanları dizi özelliklerine göredir. Tm in düşük erime alanlarına erişildiği zaman fragmentin kısımları kısmi olarak erimeye başlar, kırılan moleküller dallanma yaratmaktadır. Bu davranış DNA nın hareketini jelde azaltmaktadır. DNA fragmentindeki farklı dizilerin Tm değerleri farklılıklar (DGGE jel üzerinde farklı uzaklıklarda koşmalarına) göstermektedir. Kısacası, aynı uzunluktaki fakat farklı dizilerdeki DNA fragmentleri DGGE üzerinde ayrılabilmektedir (Ercolini, 2004). 2.1 Siyanobakteriyel Toksinler Siyanobakteriyel toksinler çok farklı kimyasal yapıdaki bileşik gruplarına sahiptir. Bunlar sitotoksinler ve biyotoksinler olarak iki gruba ayrılmıştır. Sitotoksin insanlar ve hayvanlar için öldürücü değildir, fakat bunlar nispeten algler ve memeli hücreleri için daha toksiktir. Bunlar enzimler, antibiyotikler ve antikarsinojenik faktörler yanında kimyasal yapıları çok karmaşıktır. Biyotoksinler insanlar için çok toksiktir ve hatta ölümcül etkileri olabilir. Biyotoksinler; nörotoksinler (sinir sistemini etkileyen), hepatotoksinler (karaciğeri
31 9 etkileyen) ve dermatotoksinler olarak ayrılmaktadır 2001). (Makowski and Wardas Toksin üreten siyanobakteriler kendi toksikolojik özelliklerine göre kategorize edilebilir. Bu kategorilerde nörotoksinlerden (anatoxin-a, anatoksin-a (S), saksitoksin ve neosaksitoksin); tümör promotorları (mikrosistinler ve lipopolisakkaritlerden); dermatotoksinler/tahriş edici toksin (lyngbyatoksin A, apysiatoksinler ve lipopolisakkaritler); hepatotoksinler (mikrosistin, nodularins ve cylindrospermopsin), olmasına rağmen, kimyasal yapıları temel alındığında, siklik peptidler (mikrosistin ve nodularin); nörotoksik alkaloidler (nörotoksinler ve cylindrospermopsin); ve lipopolisakkaritler olmak üzere siyanotoksinler üç ana grup içerisinde toplanmaktadır. Son zamanlarda özellikle su kaynaklarındaki geniş yayılımları dikkate alındığında çoğu bilim insanının ilgilendiği konu olan siklik peptit hepatotoksinlerin, nörotoksik alkaloidleri veya lipopolisakkaritlerden daha etkili olduğu bildirilmiştir (Çizelge 2.1) (Msagatia et al., 2006; Anjos et al., 2006) Mikrosistin a-genel yapısı, Heptapeptid hepatotoksin olan mikrosistin genellikle siyanobakterilerin birçok cinsinde (Microcystis, Anabaena, Oscillatoria, Planktothrix, Chroococcus, Nostoc, Anabaenopsis, ve Hapalosiphon) bulunmaktadır. Mikrosistin üretici tür Microcystis aeruginosa çok geniş yayılıma sahip ve değişik iklimlerde iyi gelişebilen bu organizma insan sağlığı için küresel bir tehdit oluşturmaktadır. Belirlenen bu türlerin tanımlanması ve yok edilmesini içeren bir çok araştırma gerçekleştirilmiştir (Sainis et al., 2010, Nicholson and Burch, 2001). Mikrosistin, monosiklik heptapeptid olarak tanımlanmaktadır. İçeriğinde γ-bağlı D- glutamik asit (D-Glu), D-alanin (D-Ala), β-bağlı D-erythro-bmetilaspartik asit (D-MeAsp), N- metilldehidroalanin (Mdha) ve bir tek C β- 20 amino asit, (2S,3S,8S,9S)-3-amino- -9-metoksi-2,6,8- trimetil-10-fenildeca- 4(E),6(E)-dienoik asit (Adda) diğer iki L-amino asitler değişken (X ve Z belirtilir) ve pozisyonu 2 ve 4 te halkalı yapı olarak bulunmaktadır. Değişken amino
32 10 asitlerin tek harfli bir kısaltması farklı mikrosistinleri ayırt etmek için kullanılır, örneğin en sık görülen mikrosistin içeriğindeki lösin ve arginin ile Mikrosistin-LR pozisyonunu almaktadır. Şekil 2.1 de değişiklikler gösterilmiştir (Christiansen et al., 2003; Tillett et al., 2000). Şekil 2.1: Mikrosistin ve nodularin izoformları. Amino asitlerin eksilmesi nodularin yapısı içinde kırmızı ile belirlenmiştir. Asteriks Mdha parçası Mdhb nodularin yapısı içinde değiştirilmesi ile belirlenmektedir. Kısaltmalar: Aba, amino-isobütirik asit; Dha, dehidroalanin; Mdha, N-metildehydroalanin; Met(O), metionin-s-oksit; Glu(OMe), glutamat methyl esteri; (H4)Y, 1,2,3,4- tetrahidrotyrosin; DMAdda, desmetil-adda; Dhb, dehidrobütrik asit; MeLan, N-metillantionin; ADMAdda, O-asetil-Adda (Christiansen et al., 2003). b- Biyosentezi ve genetiği, Nonribozomal olarak sentezlenen mikrotoksinler geniş multifonksiyonel enzim kompleksleri içeriğinde hem Nonribozomal peptid sentetaz (NPRS) hem de poliketit sentetaz (PKS) ile sentezlenmektedir. NRPS thiotemplate mekanizması ile peptidlerin oluşumunu hem prokaryotlarda hemde az miktarda eukaryotlarda bulunarak katalizlemektedir. NPRS lineer, siklik ve dallanmış siklik peptid sentezinden potensiyel ilaçlar olan penisilin, vancomisin ve siklosporin
, adenilasyon (A) ve peptidil taşıyıcı protein (PCP) domainlerinden oluşmaktadır.")
33 11 oluşmaktadır. NPRS modular yapıya sahiptir, her bir modul aktivasyon, tiolasyon, modifikasyon ve kondensasyonun tek özel substrat amino asitten sorumludur. Minimal modul kondenzasyon (C), adenilasyon (A) ve peptidil taşıyıcı protein (PCP) domainlerinden oluşmaktadır. Benzer şekilde PKS modulleri (PKS tip 1) multifonksiyonel megasentetaz tekrarlanan fonksiyonel birimleri organize etmektedir. PKS açil coenzim A monomerlerini bir araya getirmek için çekirdek domainleri ketosentetaz, açiltransferaz ve açil taşıyıcı proteinleri kullanmaktadır. Yapısal farklılıklar entegre edilen reaksiyonların üretilmesiyle örneğin, ketoredüksiyon, dehidrasyon veya enoly redüksiyonuyla oluşmaktadır (Pearson et al., 2010). Gen kümeleri (mcy) 55 kb uzunluğunda ve 10 adet açık okunabilen çerçevelerden (ORFs) oluşmaktadır (Şekil 2.2). mcya dan mcyc ye kadar olan kısmı beş NPRS modülleri, mcyd iki tip I PKS modulleri ve mcye ve mcyg hibrid NRPS-PKS modulerini kodlamaktadır. mcyf, mcyh, mcyi ve mcyj genleri sırasıyla racemaz, ABC transporter, D-3-fosfogliserat dehidrogenaz (D-3-PGDH) ve O-metiltransferazı (OM) kodlamaktadır. Planktothrix agardhii de bulunan mcyt geni belirgin olarak tioesteraz domaini kodlamaktadır (Pearson et al., 2010). Şekil 2.2 : Microsistin ve nodularin gen yapıları (A) Nodularia spumigena, (B) Microcystis aeruginosa,(c) Planktothrix agardhii, ve (D) Anabaena sp. 90 için gösterilmiştir. Poliketit sentetaz (beyaz), non-ribosomal peptit sentetaz (kırmızı), eklemeli enzimler (gri), ve ABCtransporterlar (siyah) olarak diyagramda açıklanmaktadır (Pearson et al., 2010).
34 12 c-toksin miktarını değiştiren parametreler, Hepatotoksin üreten siyanobakterileri azot, fosfor, iz metaller, büyüme sıcaklığı, ışık ve ph gibi farklı fiziksel ve çevresel parametrelerden etkilediği düşünülmektedir. Aslında düzenleyici araştırmaların standart hale getirilmemesi ve verilerin aynı spesifik büyüme kontrollerine karşı yorumlanmamasından dolayı hepatotoksin düzenlemesi biraz tartışmalı konu olmaya devam etmektedir. Bütün toksin düzenleme çalışmaları hücresel toksinlerin doğrudan ölçümleri üzerine yoğunlaşmış olsada, Tillett et al., (2000) tarafından mcy gen kümesinin açıklanmasında moleküler düzeyde mikrosistin düzenlemesini incelemişlerdir. Kaebernick et al., (2000) farklı ışık şiddetleri altında mcyb ve mcyd transkripsiyonu RNase koruma yöntemi ile ölçümlemişdir. Yüksek ışık şiddeti ve kırmızı ışık transkripsiyonu arttırırken, mavi ışığın azatlığı belirlenmiştir. Yazarlar gözlemlediği iki ışık eşikleri, karanlık ve düşük ışık (0 ve 16 μmol photons m -2 s -1 ) arasında ve orta ve yüksek ışıkta (31 ve 68 μmol photons m -2 s -1 ) transkripsiyonda önemli bir artış olmuştur. Ayrıca besin içeriği ve sıcak gibi diğer faktörlerin toksin biyosentezi ve mcys ekspresyonuna etkileri gösterilmiştir. Örneğin, Sevilla et al., (2008) Microcystis aeruginosa PCC7806 üzerinde demirin toksin biyosentezi ve mcys ekspresyonuna etkisini araştırmıştır. mcyd transkripsiyonu Real-time PCR da ve mikrosistin-lr sentezi HPLC kullanılarak ölçülmüştür. Bu çalışma göstermiştir ki demir eksikliği mcyd transkripsiyonunda artışa nedeniyle toksik seviyesinde de artış olmaktadır. Davis et al., (2009) Microcystis in büyümesi ve toksin genleri üzerine sıcaklığın etkilerini araştırmıştır. Sonuç olarak sıcaklığın Microcystis hücrelerinin toksisitesi ve hücre başına mcyd kopyalarının artmasına neden olduğu ve buna bağlı olarak toksik patlamanın (bloom) da arttığı gösterilmiştir Nodularin; a-genel yapısı, Nodularia spumigena ani çoğalmaları genellikle toksiktir ve bir hepatotoksin olan nodularini üretmektedirler. Nodularin siklik pentapeptid genel yapısında siklo (-D-MeAsp 1 -L-ARG 2 -Adda 3 -D-Glu 4 -Mdhb 5 ), MeAsp 1 D-eritro b- methylaspartic asit, Mdhb - (metilamino) -2 (Z)- dehidrobütirik asit
35 13 kısaltılmışıdır, Adda 5 bir (2S, 3S, 8S, 9S)-3-amino-9-metoksi-2, 6,8, -trimetil-10- phenyldeca-4 (E), 6 (E) -dienoik asit içermektedir (Moffitt and Neilan, 2001). Nodularinin yapısı ve biyolojik aktivitesi mikrosistine benzemektedir, tatlı su siyanobakterileri tarafından üretilen heptapeptid hepatotoksindir. Toksini üreten algin ani çoğalması akut durumlarda karaciğer yetmezliğine neden olduğundan suda yaşayan hayvanlar için tehlike oluşturmaktadır (Carmichael, 1994). Günümüze kadar nodularin zehirlenmesine atfedilen hiç bir insan ölümü kaydedilmemiş olmasına karşın, toksine uzun süreli maruz kalması durumunda insan sağlığı için potansiyel tehlike oluşturmaktadır (Akcalan et al., 2009). b-biyosentezi ve genetiği, Nodularin biyosentez nda genleri N. spumigena NSOR10 dizi sırası ve karakterizasyonunu Moffitt and Neilan tarafından 2004 te yapmıştır. 48 kb lık genom içeriğinde 9 ORF (ndaa-i) iki yönlü düzenleyici promotor bölgeleri transkripsiyonu gerçekleştirmektedir (Şekil 2.3). nda kodladığı genler mcy kümelerinin homologları katalitik enzimlerin kodladığı genler non ribosomal metabolit NRPS biyosentezlerinin katalitik süreçleridir. Nodularin biyosentezinin yol izi mikrosistine benzemektedir. Fonksiyonel enzimlerin tayininde biyoinformatik analiz ve mikrosistin sentetaz enzimlerinin homologunu temel almaktadır. Adda yan zincir bir fenilasetat starter ünitesinin NPRS/PKS yol izlerinin karışımına ve birkaç malonil-coa ekspresyonları (NdaC, D ve F) (Şekil 2.3) üretilmektedir (Moffitt and Neilan, 2004). NRPS ve PKS proteinleri fosfopantenil transferaz (PPT) proteini ile posttranslasyonel modifikasyonuna ihtiyaç duymaktadır. Nda proteinlerinin aktivasyonuna ihtiyaç duyan PPT diğer nda genleri ile kümelenmemiştir. Şekil 2.3: Nodularia spumigena nodularin gene yapısı (Moffitt and Neilan, 2004).
36 14 c-toksin miktarını değiştiren parametreler, nda kümesinin aydınlatılmasında biyosentetik yol izinin transkripsiyonel düzenleme izlemesi sağlamıştır. Amonyak ve fosfat kıtlığının etkileri analizlenmiştir. nda küme ekspresyonunda asıl görününen, fosfat kıtlığı sonucunda yaklaşık 2 kat ekspresyonun arttığı, amonyak eklenmesinde ekspresyonun 2 kat azaldığı belirlenmiştir. Ekspresyonun değişiklikleri, hücre içi ve hücre dışı nodularin konsantrasyonu stabil kalmıştır (Pearson et al., 2010) Anatoksinler a-genel yapısı, Günümüzde tanımlanan nörotoksik alkoloidler; anatoksin-a, homoanatoksin-a ve anatoksin-a (s) dır (Şekil 2.4). Anatoksinler sadece siyanobakterilerden Anabaena, Aphanizomenon, Cylindrospermum, Oscillatoria, Microcystis, Raphidiopsis, Planktothrix, Arthrospira, Nostoc ve Phormidium daha çok Kuzey Yarımküre tanımlanmıştır. Kanada, Amerika, İsveç, Almanya ve Afrika da yaygın olduğu yerler arasında olduğu rapor edilmiştir (Osswald et al., 2007a). Anabaena circinalis Murray-Darling havzasında Avusturalya da analiz edilmiş ve anatoksin-a ya rastlanmamıştır (Fergusson and Saint, 2000). Anatoksin-a bir alkaloiddir ve düşük molekül ağırlığına (MA=165) sahip, ikincil amin dir (Osswald et al., 2007a). Anatoksin-a hayvanlarda sinir sistemindeki nikotinik asetilkolin reseptörüne bağlanarak postsinaptik depolarizan olarak nöromuskular bloklayıcı ajan olarak görev yapmaktadır. Fakat kolinesterazı hidrolize etmez (Osswald et al., 2007b). Bu nörotoksin homolog olarak bilinen en iyi alkaloid toksin anatoksin-a içinde homoanatoksin-a da bir metil grubu eklenir ve bir hidrojen çıkması ile oluşmaktadır. Yakın zamanda toksik strain Raphidiopsis mediterranea suşundan LBRI 48 homoanatoksin-a, anatoksin-a ve yeni toksik olmayan analogu 4Shydroxyhomoanatoksin-a izolasyonu yapılmıştır (Namiksohi et al., 2004).
37 15 Anatoksin-a Homoanatoksin-a Anatoksin-a(s) Şekil 2.4 : Anatoksinlerin genel yapıları. Anatoksin-a, homoanatoksin-a ve anatoksin-a(s) (Cadel- Six et al., 2009). b- Biyosentezi ve genetiği, Gallon ve arkadaşları (1990; 1994) Anabaena flos-aquae NRC-44-1 türünden anatoksin-a biyosentezini ilk olarak araştırmıştır. Fakat bu tür anatoksina üretim kapasitesini kaybetmiştir. Diğer çalışmalarda Aphanizomenon flos-aquae IC-1 türü ile devam edilmiştir (Oswalld et al., 2007a). Oscillatoria Pasteur Culture Collection (PCC) 6506 siyanobakterisinin ürettiği anatoksin-a ve homoanatoksin-a DNA dizisi tanımlanmıştır. Bu diziye kısaca ks2 denilmektedir, ks2 dizisi PKS nin kodlandığı bilinmektedir. 29 kb DNA fragmenti içerisinde ks2 dizisi tanımlanmıştır. Genomik çerçevesinin dizisi analizlenmiş ve bulunan bazı genler Şekil 2.5 te gösterilmektedir (Mejean, 2010). Şekil 2.5 : Oscillatoria PCC 6506 anatoksin-a ve homoanatoksin-a biyosentezinden sorumlu olan tahmini gen kümelerinin organizasyonu (Mejean et al., 2010). c-toksin miktarını değiştiren parametreler, Siyanobakterilerdeki biyokimyasal veya fiziksel özelliklerinin sonuçları tam olarak belirlenmemiştir. Anatoksin-a üretiminde laboratuar koşullarındaki doğal örnekler için optimum sıcaklık 19,8-22 C arasında maksimum
38 16 konsantrasyonları 2-3 haftalık kültürlerin eksponensiyel fazında oluşmaktadır (Oswalld et al., 2007b) Saksitoksinler (Paralitik Kabuklu Zehirler (PSPs)) a- Genel yapısı, Nörotoksin saksitosinler veya paralitik kabuklu zehirleri (PSPs) denizel organizmalardan dinoflagellatların ürettiği toksinlerdir. Siyanobakteriyel türlerden saksitoksin üretiminden sorumlu tutulan üç siyanobakteri vardır. Bunlar; Aphanizomenon flos-aquae, Lyngbya wollei ve Cylindrospermopsis raciborskii dir. Danimarka göllerinde Anabaena lemmermannii nin de saksitoksin ürettiği gösterilmiştir. Toksin profilleri karmaşık ve değişkendir, dinoflagellatlar ve kontamine olan kabuklu deniz ürünlerine benzer oldukları görülmüştür (Wiese et al., 2010). Saksitoksinlerin göreceli kompleks sınıfı 18 bileşiğinin farklı olan toksisiteleri oldukça yaygın olarak üç grup altında toplanmıştır (Şekil 2.6). Bu üç grup temelde asidik koşullar altında molekülün ağ görünümün üzerinden ayrılmıştır. Bu gruplama saksitoksinler oluşur (Saksitoksin (STX), neosaksitoksin (neostx) ve decarbamoyl türevleri), gonyautoksinler (GTXs) ise decarbamoyl türevleri ve C toksinler içermektedir. PSP toksin türevleri oldukça yüksek toksisiteye (memelilerde sodyum kanalını bloke eden maddeler olarak) sahiptir (Aydın ve Uzer, 2009). Şekil 2.6: Saksitoksinlerin genel yapıları. STX = saksitoksin, NeoSTX = neosaksitoksin, GTX = Gonyautoksin (Patockaa et al. 2002).
.")
39 17 b- Biyosentezi ve genetiği, Karşıt genetik yaklaşımlarını kullanan Kellmann ve arkadaşları (2008) Cylindrospermopsis raciborskii T3 (sxt) in saksitoksinin biyosentesinden sorumlu olan tahmini gen kümeleri tanımlanmıştır. sxt gen kümeleri 35 kb dan fazla ve 26 proteinin 30 katalitik fonksiyonları karşılaştırılmalı dizi analizi ile tahmini kodlanmıştır (Şekil 2.7). Siyanobakteriyel gen kümelerinin biyoinformatik analizinde tanımlanan yeni biyosentetik yaklaşımlarla önceki geçerli olan saksitoksin biyosentez yol izinin birbiriyle bağlantısı nedeniyle geçerli olmaktadır (Pearson et al., 2010). Şekil 2.7: Paralitik Kabuklu Zehirler biyosentez yapısı (sxt); (a) Aphanizomenon sp. NH-5, (b) Anabaena circinalis AWQC131C, (c) Cylindrospermopsis raciborskii T3. Skalada gen uzunluğu kilobaz olarak belirtilmiştir (Pearson et al., 2010). c-toksin miktarını değiştiren parametreler, PSP toksinlerinin dinoflagellatlar ve siyanobakterilerde üretimini çevre faktörleri örneğin; besin (azot ve fosfat vb) içeriği, tuzluluk ve sıcaklığın düzenlediği rapor edilmiştir (Pearson et al., 2010).
40 Cylindrospermopsin a-genel yapısı, Cylindrospermopsis raciborskii izole edilen bir hepatotoksik alkaloid toksindir ve o nedenle ismi cylindrospermopsin (Şekil 2.8). Bir trisiklik alkaloid olan cylindrospermopsin bir trisiklik guanidin parçasının hidroksimetilurasil ile birleşmesiyle oluşmuştur. Belli siyanobakteriyel suşlarının doğal olarak ürettiği toksindir. Bazı suşları Cylindrospermopsis raciborskii (Avustralya, Macaristan ve ABD), Umezakia natans (Japonya) ve Aphanizomenon ovalisporum (Avustralya ve İsrail'de) cylindrospermopsin üretmediği tesbit edilmiştir (Yılmaz et al., 2008). Cylindrospermopsin toksikolojik araştırmalarda hepatotoksik alkaloid sınıfı içerisinde göstermiştir. Toksine maruz kalanlarda başlıca etkilenen organ karaciğer olup diğer etkilenen organ ise böbreklerdir (Pearson et al., 2010). Şekil 2.8: Cylindrospermopsinin genel yapısı (Clarkson, 2009). b- Biyosentezi ve genetiği, Cylindrospermopsin biyosentezi (cyr) C. raciborskii AWT205 gen kümelerinin dizileri yapılmıştır. Kümelerin genişliği 43 kb ve 15 ORF içermektedir. Bunlar bütün fonksiyonların kodlanması için toksin biyosentezi, oluşumu ve düzenlenmesinde gereklidir (Şekil 2.9).
41 19 Şekil 2.9: C. raciborskii T3 sxt gen kümelerin yapısal organizasyonu. Kısaltmalar sırasıyla: IS4, insertion sequence 4; ompr, ompr ailesinin translasyonel düzenlenmesi; hisa, iki-bileşikli histidin kinaz; orf24, ORF 4. Gen kümelerinin baz çiftlerinin uzunluğu skalda verilmiştir (Kellmann et al., 2008). c-toksin miktarını değiştiren parametreler, Cylindrospermopsin yüksek oranda gelişim ortamında azot fiksasyonun azalmasıyla gerçekleşmektedir. Cylindrospermopsin gen kümelerinin yanında hyp gen homologları hidrogenazların gelişmesini içermektedir. Nostoc PCC siyanobakterisi azot düzenleyicisi NtcA azot asimilasyonu genlerinin transripsiyonu aktive edilmektedir. Cylindrospermopsin gen kümeleri C. raciborskii AWT205 hyp gen kümeleri eksizsiz yer almasına rağmen cylindrospermopsin gen kümelerinin tanımlanmasında promotor bölge açıklanamamıştır (Pearson et al., 2010) Lipopolisakkarid (LPS) endotoksinler; Siyanobakterilerin LPS endotoksinlerin, en az ürettiği toksin olduğu anlaşılmıştır. Bu toksinler hem siyanobakterilerin ve hem de heterotrofik gramnegatif bakteriler dış duvar bileşenleridirler. Siyanobakteriler tarafından üretilen LPS endotoksinler bakteri tarafından üretilen daha az zehirli olmasına rağmen siyanobakterilerin toksinlerine maruz kalan insanlarda gastroenterit hastalıklardan sorumlu olabilir (Nicholson and Burch, 2001).
42 20 Çizelge 2.1: Siyanobakteriyel toksin grupları ve kaynakları (Codd et al.,2005). Toksin Yapısal çeşitlerinin sayısı Yapısı ve Aktivitesi Toksik türler Hepatotoksinler Mikrosistin 71 Siklik heptapeptidler; hepatoksin, protein fosfataz inhibitörü, membran bütünlüğü ve geçirgenliğin bozulması, tümor promotoru Nodularin 9 Siklik pentapeptidler; hepatotoksinler, protein fosfataz inhibisyonu, membran bütünlüğü ve geçirgenliğin bozulması, tümor promotoru, karsinojen Cylindrospermopsin 3 Guanidine alkaloidleri; Yaşamsal organlarda nekrotik zararlar (örneğin böbrekler, dalak, akciğerler, bağırsaklar), protein sentez inhibitörü, genotoksik Nörotoksinler Anatoksin-a (içerir Homoanatoksin a) 5 Alkaloidler; postsinaptik, Depolarizasyon nöromuskular bloklayıcı Anatoksin-a (s) 1 Guanidin metil fosfataz esteri; asetilkolinesterazı inhibe eder Saksitoksinler 20 Karbamat alkaloidler, Sodyum kanlarlını bloke eder Microcystis, Anabaena, Nostoc, Anabaenopsis, Planktothrix, Oscillatoria, Hapalosiphon Nodularia, Theonella (sünger-siyanobakteri ile simbiyont yaşayan) Cylindrospermopsis, Aphanizomenon, Umezakia, Anabaena, Raphidiopsis Anabaena, Oscillatoria, Phormidium, Aphanizomenon, Rhaphidiopsis Anabaena Aphanizomenon, Anabaena, Lyngbya, Cylindrospermopsis, Planktothrix Dermatotoksinler ve sitotoksinler Lyngbyatoksin-a 1 Alkaloids; inflamasyon ajanı, poretin kinaz C aktivatörü Lyngbya, Schizothrix, Oscillatoria Aplysiatoksin 2 Alkaloidler; inflamasyon ajanı, poretin kinaz C aktivatörü Endotoksinler Lipopolisakkarid Çok Lipopolisakkaritler; inflamasyon ajanı, gastrointestinal iritantlar Lyngbya, Schizothrix, Oscillatoria Hepsinde? Bu kapsamda siyanobakterilerin çok çeşitli yayılış alanlarına sahip oldukları bilinmetedir. Termal alanları ekstremofil canlılar için uygun yerlerdir ve
43 21 bu alanlar siyanobakterileri de bulundurmaktadır. Daha önce ülkemizde termal alanlarda bulunan siyanobakteriler ile ilgili bir çalışmanın mevcut olmadığı saptanmıştır. Bu nedenle yurdumuzun termal alanlarının siyanobakterilerinin belirlenmesi çalışmalarına katkıda bulunmak, bu konudaki bilgi açığının kapatılması amaçlanmıştır. İzmir ilindeki farklı termal alanlardan siyanobakteri örnekleri toplanmış, izolasyon, saflaştırma, morfolojik ve moleküler yöntemeler ile teşhisi ve toksin varlığının araştırılması amacı ile bu çalışmanın gerçekleştirilmesi planlanmıştır.
44 22 3. MATERYAL VE YÖNTEM 3.1 Organizma Bu çalışmada kullanılan siyanobakteri türlerinin orijini, İzmir ilinin farklı termal kaynaklarıdır. Seçilen istasyonlardan; Dikili-Zeytindalı ndan 1 tür, Balçova Termal alanın çıkış suyundan 2 farklı tür, Çeşme-Şifne den 1 tür, Seferihisar-Karakoç Termal alanında 2 tür, Urla-Gülbahçe termal alanından 4 farklı tür olmak üzere toplam 10 filamentli siyanobakteri türü saflaştırılıp, izole edilmiştir (Çizelge 3.1). Çizelge 3.1: İzole edilen özgün türler ve kodları İzole edilen Yer Kısaltması Dikili-Zeytindalı Zeytindalı ZD Çeşme-Şifne Şifne Şif Urla-Gülbahçe Urla 1 U sal Urla-Gülbahçe Urla 2 U ip Urla-Gülbahçe Urla 3 U pse Urla-Gülbahçe Urla 4 U osc Seferihisar-Karakoç Karakoç 1 KY Seferihisar-Karakoç Karakoç 2 KK Balçova Balçova 1 Bal ip Balçova Balçova 2 Bal pse 3.2 Kültür ortamı Siyanobakteri kültürleri uluslar arası standart metotlara uygun için Blue Green Algae (BG-11) kültür ortamı kullanılmıştır(apha, 1996). Ortam içeriği filtre edilen termal suların (Whatman No:1 filtre kâğıdı ile fitre edilerek) içinde çözüldükten sonra 121 o C de 20 dakika otoklavlanarak sterilizasyonu sağlanmıştır. Katı ortam hazırlamak için litreye 15 g agar ilave edilmiştir. Kültür ortamlarının kompozisyonu Çizelge 3.2 te verilmiştir.
45 23 Çizelge 3.2: Blue Green Algae (BG-11) ortamının kompozisyonu (Sukatar, 2004) Stoklar 1L Destile H 2 O 1 NaNO g 2 K 2 HPO 4 4 g 3 MgSO 4.7H 2 O 7,5 g 4 CaCl 2.2H 2 O 3,6 g 5 Sitrik asit 0,6 g 6 Ferik Amonyum Sülfat 0,6 g 7 Na 2 Mg EDTA 0,1 g (disodium magnesium salt) 8 Na 2 CO 3 2 g Ortam; 9 İz Metal Çözeltisi (A 5 +Co) g/1l Destile H 2 O H 3 BO 3 2,86 MnCl 2.4 H 2 O 1,81 ZnSO 4.7H 2 O 0,222 NaMoO 4.2H 2 O 0,390 CuSO 4.5H 2 O 0,08 Co(NO 3 ) 2. 6H 2 O 0,05 Stok Çözeltilerden 1-8 e kadar olanların her birinden 10 ml Stok Çözeltilerden 9 dan 1 ml 1 litrelik balon joje içine stok çözeltilerden belirtilen miktarlarda eklendikten sonra 1L ye tamamlanarak hazırlanmıştır. 3.3 Stok kültür hazırlanması Balçova, Zeytindalı, Karakoç, Şifne ve Urla termal sularına hazırlanan Blue Green Algae (BG-11) katı kültür ortamı içeren petri kabına çizgi ekim yöntemi ile inokule edilerek 10 gün boyunca koloni oluşturmak üzere özel kabinlerde inkübasyona bırakılmıştır. Bu işlem en az üç kez tekrarlanmıştır. Aynı zamanda sıvı kültürde yaşayan türler içinde seyreltme işlemi yapılmıştır. Monoalgal ve aksenik koloniler elde edilinceye kadar bu işlemlere devam edilmiştir. Daha sonra steril BG-11 sıvı ortamı içeren otoklavlanmış 10 ml lik
.")

46 24 test tüplerine ateş yanında öze yardımıyla aktarılmıştır. Bir hafta bu tüpler vorteks ile karıştırılarak çöken alglerin homojen dağılması ve havalanması sağlanmıştır. Bir hafta sonra 10 ml lik tüplerden steril 100 ml lik erlenlere aktarılarak kültür dolabında 35 C altında 15 µmol/m 2 sn ışık şiddeti altında gelişmesi sağlanmıştır (Şekil 3.1) gün Şekil 3.1: Stok kültür hazırlanması ve türlerin inkübasyon ortamları 3.4 Fiziksel koşullar Çoğaltılan örnekler 35±2º C inkübasyon kabinlerinde termostalı ısıtıcı yardımıyla inkübe edilmiştir. Bulundukları ortamın ph, ışık (24h aydınlıkta) ve sıcaklık ayarları sürekli kontrol edilmiştir. Işık kaynağı olarak Philips marka (18W/54) day light floresan lambalar kullanılmıştır.
47 Termal alandaki su özellikler Şifne Termal Alanı; İzmir in Çeşme İlçesi nin yaklaşık 5-7 km doğusundadır. Suyun ısısı 38-42ºC derecedir (Çizelge 3.3, Şekil 3.2). Çizelge 3.3: Şifne Termal Suyuna ait kimyasal kompozisyon İyonlar mg/l Potasyum K + 460,207 Sodyum Na + 8,262,146 Kalsiyum Ca +2 1,237,970 Magnezyum Mg +2 1,878,652 Demir Fe +2 0,400 Alüminyum Al +3 0,170 Çinko Zn +2 0,130 Klorür Cl 18,900,039 Bromür Br 18,900,039 Florür F 4,800 Kükürt SO -2 2,150,000 Nitrat NO -3 0,660 Hidrofosfat HPO -2 0,225 Bikarbonat HCO 3 28,600 Metaborik asit HBO 3 18,630 Metasiliat asit H 2 SiO ph: 6,76 Radyoaktivite Total (a) Acti: 106,98 Pci/L Total (a) Acti: 71,05 ± 11,16 Pci/L Radon: 850 Pci/L
 Balçova Termal Alanı; \"Agamemnon Kaplıcaları\" olarak bilinen ve 2500 yıldan beri insanlık için")
. Çizelge 3.")
48 26 Şekil 3.2: Şifne Termal Otel (Havuz) Balçova Termal Alanı; "Agamemnon Kaplıcaları" olarak bilinen ve 2500 yıldan beri insanlık için şifa kaynağı olan kaplıcalar, şimdi modern bir tesis olan Balçova Termal Tesisleri bünyesindeki Fizik Tedavi ve Rehabilitasyon Merkezi olarak hizmet vermektedir (Çizelge 3.4). Çizelge 3.4: Balçova Termal Suyuna ait kimyasal kompozisyon İyonlar mg/l Potasyum 28,4 Sodyum 375,0 Kalsiyum 16,7 Kükürt 0,1 Magnezyum 10,3 Demir 0,1 Bikarbonat 695,0 Klor 206,0 Sülfat 165,5 ph : 8,1
 Zeytindalı Termal Alanı; Bakırçay bölgesinde yer alan bu alan, termal kaynaklar")

49 27 Şekil 3.3: Balçova Termal Otel (Sıcak su çıkışı) Zeytindalı Termal Alanı; Bakırçay bölgesinde yer alan bu alan, termal kaynaklar yönünden yıllık geçmişe sahiptir(çizelge 3.5, Şekil 3.4). Çizelge 3.5: Zeytindalı Termal Suyuna ait kimyasal kompozisyon İyonlar mg/l Sodyum 219,555 Potasyum 7,038 Amonyum 0,193 Magnezyum 18,233 Kalsiyum 125,801 Mangan 0,100 Demir 1,000 Florür 2,210 Klorür 50,694 Bromür 0,150 Nitrit 0,023 Sülfat 550,000 Bikarbonat 274,500 Sülfür 0,001 Fosfat 0,090 ph 7,32
")





50 28 Şekil 3.4: Zeytindalı Termal Otel (Sıcak su çıkışı) Karakoç Termal Alanı; Alandaki aktif fayların varlığı nedeniyle jeotermal kaynak alanlarındandır. Sit Alanı olarak koruma altındadır. İzmir ili Seferihisar ilçesinin 17 km. güneydoğusunda, Kavakdere Köyü yakınında bulunan bu kaplıcanın suyu bol miktarda karbondioksit, bikarbonat ve sodyum klorür içermektedir (Şekil 3.5). Şekil 3.5: Karakoç Termal Havuzu ve sıcak su çıkışı Urla Kaplıcası; Urla ilçe merkezinin 15 km. batısında, Gülbahçe Körfezi nde deniz kenarında bulunan ılıca aynı zamanda bir hamama sahiptir (Şekil 3.6).

 Marker-Agaroz da kullanılan standart- 1 kb DNA ladder (BioLabs, N3232S) Çözeltiler;")
51 29 Şekil 3.6: Tarihi Urla-Gülbahçe Termal Havuzu 3.6 Mikroskop Saflaştırılan siyanobakterilerin morfolojik tayin yönteminin yapılabilmesi için 2 farklı mikroskopla fotoğrafları çekilmiştir. Bunlar sırasıyla; Işık Mikroskobu ve Fluoresans Mikroskobu (Leica DM 4000B) ile İYTE MAM da bulunan Scanning Elektron Mikroskobu (SEM). 3.7 Kimyasallar Marker-DGGE de kullanılan standart- 100 bp DNA ladder (BioLabs, N3231S) Marker-Agaroz da kullanılan standart- 1 kb DNA ladder (BioLabs, N3232S) Çözeltiler; Akrilamid/bis (%40) Tris-Borik Asit-EDTA (TBE) Stok solüsyon; 10X, ph:8 Tris-Asetik Asit-EDTA (TAE) Stok solüsyon; 50X Brom Fenol Mavisi Fosfat Tamponu (PBS) Çözeltisi Tris-EDTA Çözeltisi (TE) Proteinaz K % 10 SDS
52 30 %10 CTAB (Cetyl Trimethyl Ammonium Bromide) %0,7 M NaCl Fenol: Kloroform: İzoaminalkol (25:24:1) Kloroform:İzoaminalkol (24:1) 3.8 Moleküler Tanılanması a. Manuel Yöntem ile Genomik DNA İzolasyon; ml TE tamponu içerisine alınan hücreler, 5 dak. max. hızda çelik bilyeler yardımıyla vorteks edilerek hücreler patlatıldı o C de 10 dak. tutulduktan sonra hücreler tekrar 5 dak. vortekslendi rpm de 5 dak. santrifüjlenen hücrelerin üst sıvısı yeni bir ependorf tüpüne aktarıldı. 4. İçerisine 9 ml Proteinaz K ve 60 ml (%10 SDS içerisine 1 ml Mercaptoetanol) ilave edildi. 37 o C 20 dak. inkübasyona bırakıldı. 5. Üzerine 100 ml (%10 CTAB ve % 0,7M NaCl) ilave edilerek karıştırıldı ve sonra 65 o C su banyosunda 5 dak. tutuldu. 6. Toplam hacimden 600 ml alındı ve üzerine 900 ml (Fenol: Kloroform: İzoaminalkol) (25:24:1) oranında ilave edilerek ters düz edildi. 50 o C su banyosunda 5 dak. inkübe edildikten sonra rpm 5 dak. santrifüjlenen örneklerin üst kısmı yeni bir ependorf içine aktarıldı. 7. Aktarılan hacim kadar üzerine (Kloroform:İzoaminalkol) (24:1) ilave edilerek nazikçe karıştırılan tüpler santrifüjlendi ve üst sıvı yeni ependorf içine aktarıldı o C tutulanan soğuk etanol ile DNA presipitasyonu gerçekleştirildi rpm de 5 dak. santrifüjlendikten sonra pellet % 70 etanol ile yıkandı ve santrifüj sonrasında DNA kurumaya bırakıldı. 9. Kuruyan DNA ya 50 ml TE buffer ilave edilerek DNA çözüldü ve % 0,8 agaroz jel hazırlanarak elektroforoz yardımıyla görüntülendi. b. MO-BIO Bitki DNA Kiti ile Genomik DNA İzolasyon;
53 mg (0,05 g) bitki dokusu çelik bilyeli tüpe aktarılır ve üzerine 550 μl Power Plant Bead solüsyonunda ilave edilir. Vorteks yardımıyla homejanizasyonu gerçekleştirilir. 2. PB1 solüsyonu 60 o C su banyosunda kullanmadan önce ısıtılmalıdır. 3. Power Plant Bead tüpüne 60 μl PB1 soüsyonundan ilave edilir ve vorteksle karıştırılır o C su banyosunda 10 dak. inkübe edilir. 5. Max. hızla vorteks yapılarak hücrelerin parçalanması sağlanmalıdır g X 30 sn. santrifüj edilir ile 500 μl colleksiyon tüpüne alınır ve 250 μl PB2 solüsyonu ilave edilir ve karıştırıldıktan sonra 4 O C de 5 dak. inkübe edilir. 8. Tüp g de 1 dak. santrifüjlenir ve süpernatantın tamamı temiz 2.2 ml tüpe aktarılır ml PB3 ilave edilir ve 10 dak. oda sıcaklığında inkübe edilir ve g de 15 dak. santrifüj yapılır. 10. Süpernatant atılır ve pelletin üzerine 100 μl PB6 ilave edilir ve tüp havada kurutulmaz ve izopraponal ilave edilmemelidir μl PB4 solüsyonu ilave edilir ve vorteksle karıştırılır. 12. Toplam 600 μl spin filtreye aktararak g de 1 dak. santrifüj edilir. 13. Spin filtreyi yeni bir kolektör tüp içine yerleştirin ve 500 μl PB5 ilave edilip g de 30 sn santrifüjlenir. 14. Kolektör tüpündeki sıvı uzaklaştırılır ve Spin filtre temiz bir kolektör tüpüne alınır PB5 işlemi tekrarlanır. 15. Spin filtre ependorf tüpüne alınır ve üzerine 50 μl PB6 ilave edilip g de 30 sn santrifüj edilir. c. ZRFungal/Bacterial DNA Kiti ile Genomik DNA İzolasyon; mg (yaş örnek) fungus veya bakteri hücresi 200 μl su veya izotonik tampon (örneğin; PBS) ZR BashingBead Lysis Tube içerisine içerisinde çözülür ve üzerine 750 μl Lizis Solusyonu eklenmelidir. 2. Tüp max. hızda 5 dak. vortekslenir ve hücrelerin parçalanması sağlanır. 3. ZR BashingBead Lysis Tüpü 10,000 x g 1 dak. santrifüjlenir.

54 μl süpernatant Zymo-Spin IV Spin Filter (turuncu kapaklı) kolletör tüpe aktarılır ve 7,000 rpm 1 dak santrifüj yapılır. 5. 1,200 μl of Fungal/Bacterial DNA Binding Tamponu 4. basamaktaki kolektör tübe ilave edilir. 6. Zymo-Spin IIC Column kolektör tüpü içerisine yerleştirilir ve 800 μl karışımdan ilave edilir. 10,000 x g 1 dak. santrifüj edilir ve kolektör tüpündeki sıvı uzaklaştırılır ve işlem tekrarlanır. 7. Zymo-Spin IIC Column yeni kolektör tüpü içerisine yerleştirilir ve filtre üzerine 200 μl DNA Pre-Wash Tamponu ilave edilir ve 10,000 x g 1 dak santrifüjlenir ve sonra 500 μl Fungal/Bacterial DNA Wash Tamponu ilave edilir ve sonra tekrar aynı şekilde santrifüjlenir. 7. Zymo-Spin IIC Column 1,5 ml ependorf içerisine dikkatlice yerleştirilir ve üzerine 100 μl DNA Elution Tamponu ilave edilerek 10,000 x g 30 saniye santrifüjlenerek filtrede tutulan DNA toplanır. Görüntüleme; Bu üç yöntem ile elde edilen DNA lar agaroz jel elektroforezi yöntemiyle (Yatay Elektroforezi (Scie-Plas), Güç Kaynağı (Consort EV265) (Şekil 11) gözlenmiştir. Jel elektroforezi için %1,2 lik agaroz jel hazırlanmış ve örnekler 80 V, 35 ma ve 3 W da 1 saat yürütülmüştür (Çizelge 3.6, Şekil 3.7). Daha sonra etidyum bromür ile boyanan jel UV transiliminatör yardımıyla görüntülenmiştir. Şekil 3.7: Agaroz jel elektroforezi
55 33 Çizelge 3.6: Lineer DNA nın ayrılması için önerilen jel yüzdesi Agaroz Jel % Ayırma Oranı bp Poliakrilamid jel % Ayırma Oranı bp 0, , , , , , , , , , , Polimeraz Zincir Reaksiyonu (GeneAmp PCR System 9700) verilmektedir. Kullanılan primer çiftleri Çizelge 3.7, Çizelge 3.8 ve Çizelge 3.9 da Çizelge 3.7: Bakteri ve siyanobakteri belirlenmesi için seçilen primer çiftleri (*Saker et al., 2005, 1 Premanandh et al., 2006, + Green et al., 2008) Primer Hedef Populasyon 27F* Bakteri 5 -AGA GTT TGA TCC TGG CTC AG-3 809R* Bakteri 5 -GCT TCG GCA CGG CTC GGG TCG ATA R + Bakteri 5 - TAC GCG CTA CCT TGT TAC GAC-3 16S F 1 ITS Bakteri 5 - TGT GGC TGG ATC ACC TCC TT-3 23S R 1 ITS Bakteri 5 - TCT GTG TGC CTA GGT ATC CAC CGT T-3 PCβF* Siyanobakteri 5'-GGC TGC TTG TTT ACG CGA CA-3' PCαR* Siyanobakteri 5'-CCA GTA CCA CCA GCA ACT AA-3' F-1 1 Siyanobakteri 5 -TCG AAG ATC GTT GCT TGA ACG-3 R-1 1 Siyanobakteri 5 -TTA GGT CCC TGC ATT TGG GTG-3
56 34 Çizelge 3.8: DGGE de kullanılan siyanobakteri primer çiftleri (Nübel, 1997, Boutte et al.,2006) CYA 359F Siyanobakteri 5'-GGG GAA TYT TCC GCA ATG GG 3' CYA 781R(a)* Siyanobakteri 5'-GAC TAC TGG GGT ATC TAA TCC CAT T 3' CYA 781R(b)* Siyanobakteri 5'-GAC TAC AGG GGT ATC TAA TCC CTT T 3' * 40 Bazlı GC-clamp geri primere eklenmiştir; 5'-CGC CCG CCG CGC CCC GCG CCG GTC CCG CCG CCC CCG CCC G-3' Çizelge 3.8 de görüldüğü gibi CYA 781R (a) ile CYA 781(b) arasında sadece bir nükleotid farkı bulunmaktadır. Çizelge 3.9: Siyanobakteri toksinleri için seçilen primer çiftleri ( a Saker et al., 2005, b Moffitt et al., 2001, c Kellmann et al., 2008) McyA F a Mikrosistin 5 - AAA ATT AAA AGC CGT ATC AAA-3 McyA R a Mikrosistin 5 - AAA AGT GTT TTA TTA GCG GCT CAT-3 NPF b Nodularin 5 - TAT TTT GTG GTG GAG AAG CAC TA-3 NPR b Nodularin 5 - GGA ACT ATC TGA TAA TTA GAC-3 Sxt1-F c Saksitoksin 5 - GCT TAC TAC CAC GAT AGT GCT GCC G-3 Sxt1-R c Saksitoksin 5 - GGT TCG CCG CGG ACA TTA AA-3 Kullanılan 3 farklı PCR kiti; 1- Roche FastStart Taq DNA Polymerase, dntpack Ultra saf su İçerik 50 µl Final Konsantrasyon Göreceli 10x PCR Tamponu 5 µl 2mM MgCl 2 PCR Nükleotid Karışımı 1µl 200 µm (herbir dntp) İleri PCR Primer, 10X 5µl 0,2-1 µm Geri PCR Primer, 10X 5µl 0,2-1 µm Şablon DNA 5 µl FastStart Taq DNA Polimeraz 0,4 µl 2 U 27F-809R için uygulanan PCR yöntem; 50 µl ye tamamlanır Döngü Zaman Sıcaklık Denatürasyon 1 4 dak 95 ºC Denatürasyon sn 95 ºC Bağlanma ºC Uzama sn 72 ºC Son Uzama 1 7 dak 72 ºC Soğuma sınırsız 4 ºC
57 35 PCα-PCβ için uygulanan PCR yöntem; Döngü Zaman Sıcaklık Denatürasyon 1 4 dak 95 ºC Denatürasyon sn 95 ºC Bağlanma ºC Uzama sn 72 ºC Son Uzama 1 7 dak 72 ºC Soğuma sınırsız 4 ºC Roche FastStart Taq DNA Polymerase kiti kendi enziminden kaynaklanan hata nedeniyle çalıştırılamadı. O nedenle diğer 2 kite (Crimson Taq DNA Polymerase-(BioLabs) ve HelixAmpTM Speed Hifi DNA Polymerase) seçilerek kullanıldı ve 2 farklı kit ile sonuçlar başarıyla alınmıştır. 2-Crimson Taq DNA Polymerase-(BioLabs) RED ASİT içermektedir. Yöntem; İçerik 25µl 50 µl Final Konsantrasyon 5x Crimson Taq Reaksiyon Tamponu 5 µl 10 µl 1X 10 mm dntps 0,5 µl 1 µl 200 µm 10mM İleri Primer 0,5 µl 1 µl 0,2 µm (0,05-1µM) 10mM Geri Primer 0,5 µl 1 µl 0,2 µm (0,05-1µM) Şablon DNA göreceli göreceli <1000 ng Crimson Taq DNA Polimeraz 0,125 µl 0,25 µl 1,25 units/50 µl PCR Ultra pure su 25 µl 50 µl 27F-809R için uygulanan PCR yöntem; Başlangıç denatürasyon; 95 ºC 30 sn 95 ºC 20 sn 30 döngü 60 ºC 50 sn 68 ºC 59 sn
58 36 Son uzama 68 ºC 5 dak Son bekleme 4 ºC Pcα-PCβ için uygulanan PCR yöntem; Başlangıç denatürasyonu; 95 ºC 30 sn 95 ºC 20 sn 30 döngü 55 ºC 50 sn 68 ºC 59 sn Son uzama 68 ºC 5 dak Son bekleme 4 ºC 3- HelixAmp TM Speed Hifi DNA Polymerase (Nanohelix) İçerik 50 µl Final Konsantrasyon Ultra pure su Göreceli 10xReaksiyon Tamponu 5 µl 2mM MgCl 2 dntp Karıışımı 1µl 200 µm (her bir dntp) İleri PCR Primer, (10 pmoles/ml) 1~2 µl 0,2-1 µm Geri PCR Primer, (10 pmoles/ml) 1~2 µl 0,2-1 µm Şablon DNA 5 µl HelixAmp TM SpeedHifi DNA Polimeraz 0,125 µl 1,25 U 27F-809R için uygulanan PCR yöntem; Döngü Zaman Sıcaklık Denatürasyon 1 2 dak 95 ºC Denatürasyon sn 95 ºC Bağlanma 30 40sn 56 ºC Uzama sn (50 sn/kb) 72 ºC Son Uzama 1 5 dak 72 ºC Soğuma Sınırsız 4 ºC 3.10 Sekans analizi
 60 ºC de 80 V yürütülmesi a-jelin Hazırlanması 1.")
59 37 PCR sonuçlarının dizi analizi İzmir Yüksek Teknoloji Enstitüsü Biyoteknoloji ve Biyomühendislik Merkezi Araştırma Laboratuarı nda (ABI 3130XL, 16 KAPİLLAR SİSTEM) yapılmıştır. Sonuçlar NCBI-Blast programı yardımıyla değerlendirilmiştir Denature Edici Gradient Jel Elektroforez Şekil 3.8: DGGE (Scie-Plas) 60 ºC de 80 V yürütülmesi a-jelin Hazırlanması 1. Verimlilik ve poliakrilamidin çapraz bağlanmasının uniform dağılması için Electran grade materyalleri ve jel solüsyonuna degas işlemi yapılmadı. Akrilamid solüsyonu karanlıkta, soğukta örneğin buzdolabında saklandı. Isı ve güneş ışığından kaçınıldı. 2. Polimerizasyon şartları yaklaşık 15 dakikada polimerize olacak şekilde ayarlandı. Küçük hacimdeki vialde önce jel boşaltıldı. 100 ml % 6 akrilamid jel yaklaşık 5 dakikada oda sıcaklığında jelleşmesi için içerisine 450 μl taze hazırlanmış % 10 (a/h) Amonyum persülfat ve 200 μl TEMED ilave edildi. Kataliz miktarı için ılık şartlar altında yapıldı ve direkt güneş ışığına maruz bırakılmadı. 3. Jel doğrudan jel-döküm ünitesine veya sıkma tarafındaki tank birimi içine yerleştirilmiş alana döküldü.
60 38 Çizelge 3.10 da belirtilen uygun komposizyonlarda denature gradient jellerin kompozisyonları verilmektedir. Çizelge 3.10: DGGE jel kompozisyonu (farklı denature konsantrasyonları koyu renkli işaretlenmiştir) 4. Her biri 15 ml hacimlerde iki solusyon; düşük denatürant konsantrasyonu ve yüksek denatürant konsantrasyon çözeltisi hazırlandı. Gradient karıştırıcısı içerisinde solusyonların karıştırılması sağlandı ve jel gardient matriksini oluşturmaktadır. 5. APS ve TEMED içerisine ayrı ayrı ve hafifçe çalkalanarak karıştırılmaktadır. Akrilamidin polimerizasyonu başlatırlar ve yaklaşık 15 dakikada jelin katılaşmasu sağlandı. Jelin yürütülmesi; 1. Yaklaşık 5 litre 1X TAE hazırlanır ve tampon dış hazneyi kaplayacak şekilde doldurulur ve iç hazneye 0,7 litre tampon ilave edilir. DGGE aparatı içindeki tampon ön ısıtma ile 65 C ye ulaşması yaklaşık 2 saat almaktadır. 2. Sıcaklık yaklaşık 50 C kesme ısıtmaya geldiği zaman ve GRM (ana camları tutan yapı) üzerindeki jeller tampon odası içindeki yerine yerleştirilir ve kalan tampon ile iç hazneye doldurulur. 3. Her kuyu tampon ile doldurulduğunda polimerize olmayan akrilamid uzaklaştırılmaktadır. 4. Her bir kuyu tampon ile tekrar yıkanır ve yaklaşık μl boya içeren agarozda kontrol edilen PCR ürünleri kuyulara yüklenir. Yüklenen hacmin
61 39 başarısı PCR ve beklenen ürün sayısına bağlıdır. 60 C sıcaklık 20 V 10 dakika ile başlanır, sonra 200 V da 5 saat çalıştırılır (Şekil 3.8). Jelin boyanması; 1. Elektroforez tamamlandığı zaman, aparatlar ve cam plaktan jel çıkarılır. Plaklar dikkatlice ayrılarak, jel yırtılmadan çıkarılmalıdır. 2. Jel etidyum bromür ile boyanır ve DNR Bio-Imaging Systems (Chemiluminescence & Fluorescence) yardımıyla bantlar gözlenir Kompetitif=Rekabetçi ELİSA ELİSA ile mikrosistin ve nodularin belirlenmesi ticari kit (QuantiPlate TM Kit for Microcystins (Envirollogix Cat No EP022) kullanılmıştır. Örnek Hazırlanması; Stasyonel fazda hücreler ortamlarından santrifügasyon yöntemiyle uzaklaştırılmış ve liyofilizasyon işlemiyle kurutularak -20 o C de muhafaza edilmiştir. ELİSA analizi için bu hücreler sonik banyo ve sonikasyon probu ile % 75 metanol içinde parçalanmıştır. Parçalanan hücreler santrifügasyonla pelletten ayrılan süpernatat cam viallerde toplanarak 40 o C de metanolun uçurulması sağlanmıştır. Bu vialler işlem sırasında 1 mg/ml olacak şekilde ultra pure su ile çözülerek ELİSA yapılmıştır. 1. Hızlı olarak her kuyuya 125 μl Mikrosistin Analiz Seyreltici eklenir, tekrarlı ya da çok kanallı pipetter tercihen kullanılır. 2. Her biri 20 μl olarak hemen Negatif Kontrol (NC), Pozitif Kontrol (C1-C3) ve örnekler (20 μl S1-S8) kendi kuyularına eklenir (Tüm reaktifler için ek bu aynı sırayla uygulanır). Bu adımda Mikrosistin-enzim Konjugatı (Eşlenik) eklenmez. 3. Örnekler dökülmeden ve karıştırılmadan dikkatlice dairesel hareketlerle saniye sallanır. NOT: 3 ve daha fazla şeritlerin kullanımında başlatma süresini en aza indirmek için çok kanallı pipet kullanımı 1, 2, 5, 8 ve 10 uncu adımlarda tavsiye edilir.
62 40 4. İnkübasyon süresinde kuyularından örneklerin buharlaşmaması için üzeri bant veya parafilm ile kaplanarak oda sıcaklığında 30 dakika 200 rpm de çalkalanarak inkübe edilir. 5. Her kuyuya 100 μl Mikrosistin-Enzim Konjugatı eklenir adımda yapıldığı gibi kuyuların içeriği iyice karıştırılır. Kuyuları bant veya parafilm ile kapatarak 30 dakika boyunca ortam sıcaklığında çalkalamalı olarak inkübe edilir. 7. İnkübasyon sonrası dikkatlice parafilm kaplama kaldırılır ve kuvvetlice sarsılarak bir lavabo veya başka uygun bir kap içine kuyu içeriğini boşaltılır. Yıkama Çözeltisi ile kuyuları yıkanır. Bu işlemi dört defa tekrarlanır. Kuyuların içinde yıkama çözeltisi kalmadığından emin olunur. 8. Her kuyuya 100 μl Substrat ilave edilir adımda yapıldığı gibi kuyularının içeriği karıştırılır. Kuyular yeni bant veya parafilm ile kapatılarak 30 dakika boyunca ortam sıcaklığında çalkalamalı olarak inkübe edilir. 10. Her kuyuya 100 μl of Durdurma çözeltisi ilave edilir ve iyice karıştırılır. Renginin sarıya dönmesi gerekmektedir. NOT: Durdurma çözeltisinin ilavesinden sonra 30 dak. içerisinde 450 nm de Termo Scientific Multiskan FC ile okumalar yapılır. % Bo=(Ortalama Kalibratör veya Örnek OD Değeri/Ortalama Negatif Kontrol OD Değeri)*100 Kalibratör % Bo 0,16 ppb % ,60 % ,50 % Kalibratörün % Bo su hesaplandığında istenilen aralıkta olmasına dikkat edilmelidir. Aralıklar arasında çıkmayan sonuçlarda seyreltme işlemleri yapılmalıdır.
63 41 4. SONUÇLAR VE TARTIŞMA Dünya genelinde tatlı sularda yaşayan siyanobakterilere olan ilginin artması, onların birçok türünün insanlara ve diğer hayvanlara karşı ürettiği toksinleriyle zarar vermesi nedeniyledir ün üzerinde bilinen siyanobakteri türlerinden en az 30 tanesinin toksik bileşikleri olduğu bulunmuştur. Potansiyel olarak toksik türler arasında, kimyasal ve toksikolojik olarak farklı bileşikler izole edilmiş, saflaştırılmış ve karakterizasyonu yapılmıştır (Saker et al., 2005). Morfolojik olarak, siyanobakteriler 2 μm ila 40 μm çapı olan koloni ya da tek hücreli olarak filamentli veya kokkoid formlara sahip mikoorganizmlardır. Bitki ve bakteri ikili özellikleri nedeniyle, taksonomik sınıflandırmada botanikçiler ve bakteriyologlar arasında polemik olmuştur. Klasik taksonominin temeli organizmaların morfolojik özellikleri dayanmasıdır ancak bu organizmaların çevresel koşullar altında morfolojisini değiştirebilme özelliklerinden dolayı bu sınıflandırma şeklinin son yıllarda eksik olduğu düşünülmektedir. Drouet (1968) aynı fenotiplerde oluşan genetik değişkenliğe dikkat çekmiştir. Diğer bakteriler içerisinde, günümüzdeki polifazik yaklaşımı nedeniyle taksonomik sınıflandırma en istenir halindedir. Bergey's Manual nin Sistematik Bakteriyoloji kitabının son baskısında Siyanobakteri phylumu bakteri domaini içerisinde göstermiştir. Taksonların terminolojisi Botanik Kodlarla adlandırılması yerine bakteriyolojik koduna çevrilmiştir ve Ripkka et al., (1979) tarafından aşağıdaki ana grubu önermiştir. I -tek hücreli siyanobakteriler ikili fizyon veya tomurcuklanma ile çoğalanlar, II-tek hücreli siyanobakteriler birden fazla fisyon ile çoğalanlar, III - filamentli olup heterosist içermeyenler siyanobakteriler sadece bir düzlemde bölünenler, IV- filamentli heterosist içeren siyanobakteriler sadece bir düzlemde bölünenler, V- filamentli heterosist içeren siyanobakteriler bir düzlemde daha çok bölünenler (Osswald et al., 2007b). Desikachary (1959) a göre mavi-yeşil algler bir sınıf ve 5 Ordo altında toplanmıştır (I: Ordo Chroococcales, II: Ordo Pleurocapsales, III: Ordo Chamaesiphonales, IV: Ordo Nostocales, V: Ordo Stigonematales).
64 42 Siyanobakterilerin filogenetik analizleri 16S rdna dayalı olarak çeşitli araştırma grupları tarafından yapılmış ve Chroococcales (I) ve Chamaesiphonales (III) in polifiletik (birden fazla ata türden türemiş) olduğu belirlenmiştir yılında Pleurocapsales in monofiletik (II) olduğunu reddedilmiş, heterosistik formlar olan Nostocales (IV) ve Stigonematales (V) in ise monofiletik olduğu saptanmıştır (Korelusova, 2008). Örneklerin izolasyonlarında değişik saflaştırma yöntemleri kullanılmıştır. Seyreltme yöntemi tekniğiyle Urla türleri başarılı bir şekilde ayrılmasının sağlanması için aynı zamanda fiziksel koşullardan (farklı ışık şiddetlerinde) da yararlanılmıştır (Sukatar, 2004). Seferihisar-Karakoç türleri için agarlı katı ortam hazırlanmış ve agardan tek filamentlerin steril tek kullanımlık özeler yardımıyla seçilmiş ve yeni agarlı ortamlara aktarılmıştır. Birbiri içine giren ipliksi türlerin ayrımında kısa süreli sonik banyo kullanılarak sonikasyonla ayrılmaları sağlanmaya çalışılmış ve sonra agar plağı üzerine yayılan sıvıdan ayrılmaları için inkübasyona bırakılmıştır. Zeytindalı ve Şifne ortamlarından alınan organizmalarda ise baskın duruma geçen filamentler ortamdan mikroskop altında toplanarak seçilmiştir. Hücrelere havalandırma veya çalkalama uygulanmamıştır. BG-11 Ortamı distile su yerine filtre edilmiş kendi termal sularına hazırlanmasıyla seçici ortamları hazırlanmıştır. Hareketli veya hareketsiz hormogonyuma sahip olan filamentli cinsler (örn. Oscillatoria, Phormidium, Lyngbya, Symploca, Calothrix, Mastigocladus) genellikle, uygun sıcaklıkta inkübe edilmiş ve %1,0-1,5 lik agarlı D ortamı ve Cg-10 ortamında birkaç saat içinde kayma yolu ile kolayca izole edilebildikleri belirtilmiştir (Castenholz, 1969). Tek hücreli bir alg olan M. concinna; cool-white floresans lambası altında ve Medium D ortamı içinde kültürü yapılmıştır. Kültür; tüm gün boyunca 45 ºC de ve havalandırmalı olarak korunmuştur (Sun and Wang, 2003). Halomicronema excentricum için deniz suyu ortamına benzer olması için gerekli besinler vitaminler ve iz elementler eklenerek ortam (1/2 PES) hazırlanmıştır. Mikroskop teknikleri morfolojik açıdan mikroorganizmaların çeşitliliğinin belirlenmesi, ekolojisi ve fizyolojik adaptasyonları için en sık olarak kullanılan
65 43 tekniklerdendir. Örneğin klasik ışık mikroskobunun yanı sıra scanning elektron mikroskobu (SEM), transmisyon elektron mikroskobu (TEM) ve konfokal lazer tarama tarama kullanılan mikroskobu (CLSM) kullanılmaktadır (Marine et al., 2004). Fluoresans Mikroskobunda canlı hücreler görüntülenerek siyanobakterilere özel fikobiliproteinlerinin (fikosiyanin ve fikoeritrin) fluorasans verme özelliğinden yararlanılmıştır (Şekil b). Fikosiyanin ve fikoeritrin güçlü floresan yayabilmektedir, iyi absorpsiyon performans ve yüksek kuantum verimliliğine sahiptir, bunların floresan gücü flöresein 30 katıdır ve görünür spektrumu geniş bir uyarma ve emisyon aralığında sahiptir (Liu, 2010). Görüntüleme için ayrıca bir boyama tekniği kullanılmamıştır. Mikroskop kullanımı ile elde edilen hücrelerin yeşil alg olmadıkları siyanobakteri oldukları doğrulanmıştır. SEM organizmaların yüzey topolojisi ve örneklerinin dağıtımlarını, daha doğrusu yüksek büyütmede organizmalar arasındaki ilişkiyi incelemek için kullanılmaktadır. Konvansiyonel SEM örnek hazırlama teknikleri; hava kurumalı, kimyasal fiksasyonun izlenmesinde kritik noktada kurutma veya donma fiksasyonu dondurularak-kurutma ve kritik nokta kurutmasıdır (Marine et al., 2004). Biyolojik materyalin iletkenliğinin ağır metal tuzlarıyla bütün malzemenin doyurulmasıyla arttırılmaktadır. Daha sonra numuneler taslakların üzerine yerleştirilir ve karbon veya altınla kaplama püskürtülür. Örneklerin çökme eğilimlerinde (sıvı ortam içerisinde) ön fiksasyon için akrolein buharına ek olarak tetroxid buharı verilir. Sonuçta üç boyutlu yapıları yeterli olarak korunmaktadır (Marine et al., 2004). Temiz örnekler veya kalkerli yapılı hücrelerle çalışılmış ve prosedürün özellikle kılıfları üzerindeki kalsit kristalleri veya bir dış yüzeyde depolandığını gösterilmiştir (Şekil 4.2, 4.3, 4.7, 4.9). Hücreler zarar görmemesi için kendi ortamlarında saf su ile yıkama işlemi gerçekleştirilmeden liyofilize edilmiştir. Bu yöntemin seçilmesinde, siyanobakterilerin biyolojik maddelerle dehidratasyona



.")
")

66 44 duyarlı olma eğilimindendir. Kılıfları ve müsilajlı dış katmanları yoğun veya örneklerinin yüzeyinde bulanık olarak görülmektedir (Şekil 4.3, 4.4). a b c Şekil 4.1: Urla 1 (U sal) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning Elektron mikroskobu

")




67 45 a b c Şekil 4.2: Karakoç 1 (KY) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu

")




68 46 a b c Şekil 4.3: Karakoç 2 (KK) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu


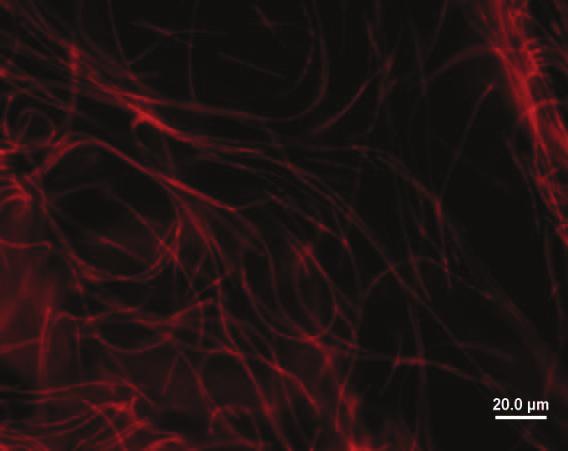



69 47 a b c Şekil 4.4 : Balçova 1 (Bal ip) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu

")



70 48 a b c Şekil 4.5: Şifne (Şif) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu

")



71 49 a b c Şekil 4.6: Balçova 2 (Bal pse) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu






72 50 a b c Şekil 4.7: Zeytindalı (ZD) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu

")



73 51 a b c Şekil 4.8: Urla 2 (U ip) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu

")



74 52 a b c Şekil 4.9: Urla 3 (U pse) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu
")





75 53 a b c Şekil 4.10: Urla 4 (U osc) fotoğrafları a. Işık mikroskobu, b. Flouresans mikroskobu, c. Scanning elektron mikroskobu Örnekler, ışık, fluoresans ve scanning elektron mikroskopta (SEM) incelenmiştir. Morfolojik yapılarına göre incelendiğinde örneklerin ortak şekilde ipliksi, çok ince kılıflı ya da kılıfsız, gerçek dallanma göstermeyen, yalancı
76 54 dallanma gösterebildiği, trikomun organizasyonlu, heterosistsiz, hormogoniumun mevcut, uca doğru karakteristik özellik gösteren yapıdadır. Bu özellikler temel alındığında bu örneklerin Nostocales ordosunda bulunduğu düşünülmüştür (Şekil ). Divisio : Cyanophyta Classis : Cyanophyceae Ordo : Nostocales Family : Oscillatoriaceae (Desikachary, 1959) Siyanobakteriler subsection III (Filamentli dahil oldukları), neredeyse tüm filamentli siyanobakterin yer aldığı aynı düzlemde ikili bölünme yeteneğine sahip olup heterosist içermeyen yapılar bulunmaktadır. Oscillatoria cinsi yaygın olarak taze, deniz ve acısularda dağılım göstermektedir. Bazı türler neredeyse siyah ve klorofil, fikoeritrin ve fikosiyanin dışında kromatik adaptasyon gösterebilmektedir (Şekil 4). Bazı mikrobiyal kümelerinin hareketli parçaları olduğu bilinmektedir (Rasoulouiriana et al., 2009). Geleneksel yaklaşım morfoloji ve ekolojiye dayalıdır, onun en önemli temsilcisi olarak Geitler (1932) gösterilmektedir. Geitler in yaptığı sistem, az veya fazla belirsizlikle takson tanımlamasıdır, nispeten kolaylık sağlamasından dolayı kullanımı hala yaygındır. Özellikle sahadan toplanan doğal örneklerin içinde siyanobakteriler belirlenmesinde fikologlar tarafından tercih edilmektedir. Ancak, Geitler (1932) in geleneksel yaklaşım sisteminde tek başına türler bildirilmekte, taksonlar arasındaki evrimsel ilişkiler yansıtılmamaktadır. Siyanobakterin morfolojik çeşitliliğinde farklı çevre koşullarının sınırlı sayıda genotiplerin üzerinde etkili olduğu sonucunu kabul eden, tamamen farklı bir anlayış Droulet (1968) tarafından önerilmiştir. Bu nedenle, bu görüşün cins sayısını önemli ölçüde azalttığı, fakat böylece mevcut genetik değişkenliği eksik değerlendirdiği ortaya çıkmıştır (Lokmer, 2007). Siyanobakteri sistematiğininde durum hala çok karmaşıktır (Komarek, 2006). Castenholz ve Phylum (2001) sistemi bakteriyolojik kodu temel alırken Komarek ve Kastovsky (2003) sistemi botanik kodlu isimlendirme kurallarına göre oluşturmuştur. Hem bakteriyologlar ve hem de botanikçiler tarafından ortak kabul edilebilir isimlendirme yapılması Oren (2004) tarafından önerilmiştir.
77 55 Problemin en kötü yanı, birçok araştırıcının resmi isimlendirmedeki herhangi bir kodu tanımlamamasıdır. Suşlar literatürde düzeltilmemiş ve genellikle doğru olmayan taksonomik tanılamalar yapılmıştır (Lokmer, 2007). Mavi-yeşil alg olarak bakıldığında Nostocales ordosu altındaki Oscillatoriaceae ailesine ait olduğunu belirlediğimiz örnekler, sınıf ve cins düzeyinde morfolojik olarak tayin edilememiştir. Siyanobakterilerin 16S ribozom içermesi ve prokaryotik olması nedeniyle örneklerin moleküler yöntemlerle tür tayini yapılmasına karar verilmiştir. Ulcay et al., 2007 tarafından yapılan çalışmada Dikili ve çevresinde seçilen 4 kaplıcada toplam 7 istasyon belirlemiş ve örneklemeler yapılmıştır. Cyanophyceae sınıfından Chroococcales ordosunda 2 tür ve Oscillatoriales ordosunda 17 türü morfolojik olarak belirlemişlerdir. Termofillerin özgün türlerinin çok geniş bir coğrafik dağılım gösterdiği gözlenmiştir. Bununla birlikte; bu konuyla ilgili kategorik bir anlatım yapmak zordur. Öncelikle siyanobakterilerin türe özgü tanımlanması pek mümkün değildir (Castenholz, 1969). Çalışmalar; çevresel örneklemelerden elde edilen 16S rrna gen bilgisinden gözlem ve kültür tekniklerinden tahmin edildiği gibi genotipik çeşitliliğin fenotipik çeşitliliğe göre çok üstün olduğunu göstermek için yararlanmışlardır (Ferris et al., 1996). Siyanobakteriyel DNA izolasyonu ve saflaştırılmasında en genel olarak lizozim, Proteinaz K, SDS, CTAB, fenol: kloroform: izoaminalkol (25:24:1) kullanılmıştır (Saha et al., 2005; Hongmei et al., 2005; Fiore et al., 2000; Adams, 1988; Rodriguez et al., 2007; Neilan, 2002). Kütle örneklerinden DNA nın elde edilmesi CTAB içinde lizozim, RNAse A ve Proteinaz K kullanımıyla, 60 0 C de fenol: kloroform ekstraksiyonu ile sağlanmıştır (Hongmei et al., 2005). Siyanobakteriyel kültür örneklerinin total genomik DNA izolasyonu sukroz tamponu içinde lizozim ve proteinaz K, SDS (sodyum dodesil sülfat) kullanımıyla fenol:kloroform:izoamilalkol ekstraksiyonuyla gerçekleştirilmiştir (Lu et al., 1997).
.")
 kullanılarak elde edilmiştir.")

, 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0 kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb); 1, ZD; 2, Şif; 3, Bal ip; 4, Bal pse; 5, U sal;")
78 56 Saflaştırılan siyanobakteri türlerin DNA izolasyonları manuel, bakteri/fungal DNA izolasyon kiti ve Mo-Bio Bitki Kiti DNA kiti kullanılarak elde edilmiştir. Elde edilen DNA lar % 0,8 lik agaroz jel de, etidyum bromürle boyamanmasından sonra görüntülenerek karşılaştırılmıştır (Şekil 4.11). Elde edilen en iyi sonuçlar bakteri/fungal DNA kitinden elde edilmesiyle bu kitin kullanılmasına karar verilmiştir. Tüm türler için DNA izolasyonları Zymo Bakteri/fungal DNA izolasyon kiti (Şekil 4.12) kullanılarak elde edilmiştir. M a M b c Şekil 4.11: Agaroz jel üzerinde kitlerden elde edilen DNA ların karşılaştırılması. a.mo-bio Bitki Kiti kullanılarak elde edilenler, b. ZRFungal/Bacterial DNA Kiti kullanılarak elde edilenler, c. Manuel Yöntem ile izole edilen DNA örneklerinin görüntüsüdür. M Şekil 4. 12: ZRFungal/Bacterial DNA Kiti ile DNA izolasyonu. Marker(M), 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0 kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb); 1, ZD; 2, Şif; 3, Bal ip; 4, Bal pse; 5, U sal; 6, U ip; 7, U pse; 8, U osc; 9, KK; 10, KY DNA elde edilmesinden sonra, bakteri universal primer ve siyanobakteri fikosiyanin gen bölgeleri için kullanılan primer çiftleri ile PCR yapılmıştır. 3

79 57 farklı PCR kiti kullanılmıştır. İlk kullanılan Roche (FastStart Taq DNA Polymerase, dntpack) marka kitin içerisinde bulunan DNA Polimeraz enziminin çalışmaması nedeniyle sonuç alınamamıştır. Bu nedenle farklı kitler sipariş edilerek elde edilen sonuçlar karşılaştırılarak doğrulanmıştır. 25 µl olarak hazırlanan PCR ürünlerinde 27F-809R için annealing sıcaklığı 62 ºC, 16S-23S için Nanohelix kitinde 65 ºC ve Crimson kiti için 67 ºC, 27F- 1492R için 56 ºC derece kullanılarak optimizasyonları yapılmıştır (Şekil 4.13, 4.14, 4.15, 4.16, 4.17, 4.18). Türler farklı olduğu için bağlanma sıcaklıkları her zaman aynı olmamaktadır, Şekil 17 ve 21 de fazla bantların oluşabildiği gözlenmektedir. Bağlanma sıcaklığın 1-2 ºC lik oynanma ile tek bant görünümleri elde edilmiştir. 3, M 0 Şekil 4.13: Nanohelix kiti kullanılarak 16SF-23SR bölgesinin çoğaltılan görüntüsü. Marker(M); 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0 kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb), 1. Sif, 2. KY, 3. Bal pse, 4. U pse, 5. KY, 6. ZD, 7. U osc, 8. U sal, 9. Bal ip, 10. KK, M
; 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0 kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb), 1. U sal, 2. Bal ip, 3. KY, 4.")

80 M Şekil 4.14: Crimson kiti kullanılarak 16SF-23SR bölgesinin çoğaltılan görüntüsü. Marker(M); 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0 kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb), 1. U sal, 2. Bal ip, 3. KY, 4. U osc, 5. U pse, 6. Bal pse, 7. KK, 8. Sif, 9. KK, 10. ZD, M M Şekil 4.15: Nanohelix hypersense kiti kullanılarak 27F-809R bölgesinin çoğaltılan görüntüsü. Marker(M); 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb), 1. KK, 2. U sal, 3. Sif, 4. Bal pse, 4. ZD, 5. Bal ip, M
; 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb), 1. U pse, 2. Bal pse, 3.")
; 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb), 1. U pse, 2. Bal pse, 3.")

81 M Şekil 4.16: Nanohelix hypersense kiti kullanılarak 16SF-23SR bölgesinin çoğaltılan görüntüsü. Marker(M); 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb), 1. U pse, 2. Bal pse, 3. Bal ip, 4. U osc, 5. KK M Şekil 4.17: Nanohelix hypersense PcβF-PCαR bölgesinin çoğaltılan görüntüsü. Marker (M); 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb), 1. U pse, 2. Bal pse, 3. Bal ip, 4. U osc, 5. KK, 6. KY, 7. U ip, 8. ZD. 5 5 M Şekil 4.18: Nanohelix hypersense 27F-1492R bölgesinin çoğaltılan görüntüsü. Marker(M); 1kb DNA ladder (10,0 kb, 8 kb, 6,0 kb, 5,0 kb, 4,0kb, 3,0 kb, 2,0 kb, 1,5 kb, 1,0 kb ve 0,5 kb), 1. U pse, 2. Bal pse, 3. Bal ip, 4. U osc, 5. KK, 6. KY, 7. U ip.
82 60 Dizi analizi sonuçları NCBI-Blast programından yararlanılarak yapılmış, ileri ve geri primer yönleri ayrı ayrı okutularak karşılaştırmalı olarak sonuçlar verilmiştir. Çıkan sonuçlar Çizelge 4.1, 4.2, 4.3, 4.4, 4.5 karşılaştırmalı olarak verilmiştir. Çizelge 4.1: 27F-809R için dizi analizi sonuçları Primerler Organizma Adı Maksimum Tanımlanma 1 AF (27F- 809R) U pse Oscillatoria rosea IAM M-220 gene for 16S ribosomal RNA % 98 1 AR (27F- 809R) U pse Oscillatoria rosea IAM M-220 gene for 16S ribosomal RNA % 98 2 AF (27F- 809R) U osc Uncultured cyanobacterium clone UFDCBA_30f 16S ribosomal % 97 RNA gene, partial sequence 2 AR (27F- 809R) U osc Uncultured cyanobacterium clone UFDCBA_30f 16S ribosomal % 96 RNA gen 3 AF (27F- 809R) KK Lyngbya birgei CCC S ribosomal RNA gene, partial %99 sequence 3 AR (27F- 809R) KK Lyngbya birgei CCC S ribosomal RNA gene, partial % 84 sequence 4 AF (27F-809R) Bal pse Uncultured cyanobacterium clone Mat-CYANO-S19 16S % 97 ribosomal RNA gene, partial sequence 4 AR (27F-809R) Bal pse Uncultured cyanobacterium clone Mat-CYANO-S19 16S % 96 ribosomal RNA gene, partial sequence 5 AF (27F- 809R) ZD Geitlerinema sp. CR_13M 16S ribosomal RNA gene, partial % 96 sequence 5 AR (27F- 809R) ZD Geitlerinema sp. CR_13M 16S ribosomal RNA gene, partial % 96 sequence 6 AF (27F- 809R) Şif Geitlerinema sp. PCC 7105 small subunit ribosomal RNA gene, % 99 partial sequence 6 AR (27F- 809R) Şif Geitlerinema sp. PCC 7105 small subunit ribosomal RNA gene, % 100 partial sequence 7 AF (27F- 809R) Bal ip Geitlerinema sp. PCC S rrna gene (partial), trna-ile % 99 gene, trna-ala gene and ITS1 and 23S rrna gene (partial), strain PCC AR (27F- 809R) Bal ip Geitlerinema sp. PCC S rrna gene (partial), trna-ile % 99 gene, trna-ala gene and ITS1 and 23S rrna gene (partial), strain PCC AF (27F- 809R) KY Phormidium animale PMC partial 16S rrna gene, strain % 99 PMC AR (27F- 809R) KY Phormidium animale PMC partial 16S rrna gene, strain % 98 PMC AF (27F- 809R)U sal Halospirulina sp.'ccc Baja-95 Cl.3', 16S rrna gene % 94 9 AR (27F- 809R) U sal Halospirulina sp.'ccc Baja-95 Cl.3', 16S rrna gene % AF (27F- 809R) U ip Geitlerinema sp. CCY S ribosomal RNA gene, partial %92 sequence 10 AR (27F- 809R) U ip Geitlerinema sp. CCY S ribosomal RNA gene, partial sequence %93 U pse 27F-809R Oscillatoria rosea IAM M-220 gene for 16S ribosomal RNA Length=1436 Score = 1306 bits (707), Expect = 0.0 Identities = 728/737 (98%), Gaps = 5/737 (0%) Strand=Plus/Plus Query 18 TGC-AGTCG-ACG-AGTCTTCGGACTTAGTGGCGGACGGGTGAGTAACGCGTGAGAATCT 74 Sbjct 28 TGCAAGTCGAACGAAGTCTTCGGACTTAGTGGCGGACGGGTGAGTAACGCGTGAGAATCT 87 Query 75 GCCTTCAGGATGGGGACAACAGTTGGAAACGACTGCTAATACCCAATATGCCTACGGGTG 134 Sbjct 88 GCCTTCAGGATGGGGACAACAGTTGGAAACGACTGCTAATACCCAATATGCCTACGGGTG 147
83 61 Query 135 AAACTTATATGGCCTGAAGATGAGCTCGCGTCTGATTAGCTAGTTGGTAGTGTAAGGGAC 194 Sbjct 148 AAACTTATTTGGCCTGAAGATGAGCTCGCGTCTGATTAGCTAGTTGGTGGTGTAAGAGAC 207 Query 195 TACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGAC 254 Sbjct 208 TACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGAC 267 Query 255 ACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTG 314 Sbjct 268 ACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAG-CTG 326 Query 315 ACGGAGCAATACCGCGTGAGGGAGGAAGGTTTTTGGATTGTAAACCTCTTTTCTCAGGGA 374 Sbjct 327 ACGGAGCAATACCGCGTGAGGGAGGAAGGTTTTTGGATTGTAAACCTCTTTTCTCAGGGA 386 Query 375 AGAATTTTGACGGTACCTGAGGAATAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTA 434 Sbjct 387 AGAATTTTGACGGTACCTGAGGAATAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTA 446 Query 435 ATACGGAGGGTGCAAGCGTTATCCGGAATTATTGGGCGTAAAGAGTCCGTAGGTGGTCAT 494 Sbjct 447 ATACGGAGGGTGCAAGCGTTATCCGGAATTATTGGGCGTAAAGAGTCCGTAGGTGGTCAT 506 Query 495 GCAAGTCTGCTGTCAAAGCCCACAGCTTAACTGTGGATCGGCGGTGGAAACTGCATGACT 554 Sbjct 507 GCAAGTCTGCTGTCAAAGCCCACAGCTTAACTGTGGATAGGCGGTGGAAACTGCATGACT 566 Query 555 TGAGTACGGTAGGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTGGGA 614 Sbjct 567 TGAGTACGGTAGGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTGG-A 625 Query 615 AGAACACCGGTGGCGAAAGCGCTCTACTGGACCGTAACTGACACTGAGGGACGAAAGCCA 674 Sbjct 626 AGAACACCGGTGGCGAAAGCGCTCTACTGGACCGTAACTGACACTGAGGGACGAAAGCCA 685 Query 675 AGGTAGCGAAAGGGATTAGATACCCCTGTAGTCTTGGCCGTAAACGATGGATACTAGGCG 734 Sbjct 686 AGGTAGCGAAAGGGATTAGATACCCCTGTAGTCTTGGCCGTAAACGATGGATACTAGGCG 745 Query 735 TTGTCTGTATCGACCCG 751 Sbjct 746 TTGTCTGTATCGACCCG 762 R Oscillatoria rosea IAM M-220 gene for 16S ribosomal RNA Length=1436 Score = 1295 bits (701), Expect = 0.0 Identities = 726/737 (98%), Gaps = 6/737 (0%) Strand=Plus/Minus Query 10 CATCG-TTACGGTC-AGACTACA-GGGTATCTAATCCCTTTCGCTACCTTGGCTTTCGTC 66 Sbjct 734 CATCGTTTACGGCCAAGACTACAGGGGTATCTAATCCCTTTCGCTACCTTGGCTTTCGTC 675 Query 67 CCTCAGTGTCAGTTACGGTCCAGTAGAGCGCTTTCGCCACCGGTGTTCTTCCCAATATCT 126 Sbjct 674 CCTCAGTGTCAGTTACGGTCCAGTAGAGCGCTTTCGCCACCGGTGTTCTT-CCAATATCT 616 Query 127 ACGCATTTCACCGCTACACTGGGAATTCCCTCTACCCCTACCGTACTCAAGTCATGCAGT 186 Sbjct 615 ACGCATTTCACCGCTACACTGGGAATTCCCTCTACCCCTACCGTACTCAAGTCATGCAGT 556 Query 187 TTCCACCGCCGATCCACAGTTAAGCTGTGGGCTTTGACAGCAGACTTGCATGACCACCTA 246 Sbjct 555 TTCCACCGCCTATCCACAGTTAAGCTGTGGGCTTTGACAGCAGACTTGCATGACCACCTA 496 Query 247 CGGACTCTTTACGCCCAATAATTCCGGATAACGCTTGCACCCTCCGTATTACCGCGGCTG 306
84 62 Sbjct 495 CGGACTCTTTACGCCCAATAATTCCGGATAACGCTTGCACCCTCCGTATTACCGCGGCTG 436 Query 307 CTGGCACGGAGTTAGCCGGTGCTTATTCCTCAGGTACCGTCAAAATTCTTCCCTGAGAAA 366 Sbjct 435 CTGGCACGGAGTTAGCCGGTGCTTATTCCTCAGGTACCGTCAAAATTCTTCCCTGAGAAA 376 Query 367 AGAGGTTTACAATCCAAAAACCTTCCTCCCTCACGCGGTATTGCTCCGTCAGGCTTTCGC 426 Sbjct 375 AGAGGTTTACAATCCAAAAACCTTCCTCCCTCACGCGGTATTGCTCCGTCAG-CTTTCGC 317 Query 427 CCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCC 486 Sbjct 316 CCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCC 257 Query 487 AGTGTGGCTGCTCATCCTCTCAGACCAGCTACTGATCGTCGCCTTGGTAGTCCCTTACAC 546 Sbjct 256 AGTGTGGCTGCTCATCCTCTCAGACCAGCTACTGATCGTCGCCTTGGTAGTCTCTTACAC 197 Query 547 TACCAACTAGCTAATCAGACGCGAGCTCATCTTCAGGCCATATAAGTTTCACCCGTAGGC 606 Sbjct 196 CACCAACTAGCTAATCAGACGCGAGCTCATCTTCAGGCCAAATAAGTTTCACCCGTAGGC 137 Query 607 ATATTGGGTATTAGCAGTCGTTTCCAACTGTTGTCCCCATCCTGAAGGCAGATTCTCACG 666 Sbjct 136 ATATTGGGTATTAGCAGTCGTTTCCAACTGTTGTCCCCATCCTGAAGGCAGATTCTCACG 77 Query 667 CGTTACTCACCCGTCCGCCACTAAGTCCGAAGACTTCGTTCGACTTGCATGTGTTAGGCA 726 Sbjct 76 CGTTACTCACCCGTCCGCCACTAAGTCCGAAGACTTCGTTCGACTTGCATGTGTTAG-CA 18 Query 727 TACCGCCAGCGTTCATC 743 Sbjct 17 TACCGCCAGCGTTCATC Synechococcus sp. PCC 7117 AB Synechococcus sp. PCC 8807 AF Rhabdoderma cf. rubrum kopara-ch AJ62... Synechococcus elongatus CCMP1630 AY94... Upse 27F-809R Oscillatoria rosea IAM M-220 AB Uncultured Oscillatoria sp. EF Oscillatoria sp. CCAP 1459/13 AY Phabdoderma sp. MBIC10210 AB Phormidium sp. UIO 146 FM U osc Uncultured cyanobacterium clone UFDCBA_30f 16S ribosomal RNA gene, partial sequence Length=1454 Score = 1266 bits (685), Expect = 0.0 Identities = 718/734 (97%), Gaps = 1/734 (0%) Strand=Plus/Plus Query 29 AGTCGCACG-ACTCTTCGGAGTGAGTGGCGGACGGGTGAGTAACGCGTGAGAATCTGCCT 87 Sbjct 54 AGTCGAACGAACTCTTCGGAGTGAGTGGCGGACGGGTGAGTAACGCGTGAGAATCTGCCT 113 Query 88 GTGGGTCGGGGATAACCACTGGAAACGGTGGCTAATACCGGATGAGCCGAAAGGTAAAAG 147 Sbjct 114 GTGGGTCGGGGACAACTACTGGAAACGGTGGCTAATACCGGATGAGCCGAAAGGTAAAAG 173
85 63 Query 148 ATTTATTGCCCACAGATGAGCTCGCGTCTGATTAGCTAGTTGGTGTGGTAAAAGCGCACC 207 Sbjct 174 ATTTATTGCCCACAGATGAGCTCGCGTCTGATTAGCTAGTTGGTAAGGTAAAAGCGTACC 233 Query 208 AAGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGG 267 Sbjct 234 AAGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGG 293 Query 268 CCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGG 327 Sbjct 294 CCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGG 353 Query 328 AGCAAGACCGCGTGGGGGAGGAAGGTTTTTGGATCGTAAACCCCTTTTGTCAAGGAAGAA 387 Sbjct 354 AGCAAGACCGCGTGGGGGAGGAAGGTTTTTGGATCGTAAACCCCTTTTGTCAAGGAAGAA 413 Query 388 GCTCTGACGGTACTTGACGAATAAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATA 447 Sbjct 414 GCTCTGACGGTACTTGACGAATAAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATA 473 Query 448 CGGAGGAGGCAAGCGTTATCCGGAATAATTGGGCGTAAAGAGTCTGTAGGTGGCGAACTA 507 Sbjct 474 CGGAGGAGGCAAGCGTTATCCGGAATGATTGGGCGTAAAGCGTCTGTAGGTGGCCAGCTA 533 Query 508 AGTCTGCTGTTAAAGATCACAGCTCAACTGTGGGTAGGCAGTGGAAACTGGTTAGCTAGA 567 Sbjct 534 AGTCTGCTGTTAAAGATCACAGCTCAACTGTGGGAAGGCAGTGGAAACTGGCAAGCTAGA 593 Query 568 GGCTGGTAGGGGTAAAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGATATCGGGAAGA 627 Sbjct 594 GGTTGGTAGGGGTAAAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGATATCGGGAAGA 653 Query 628 ACACCGGTGGCGAAAGCGCTTTACTGGGCCAGACCTGACACTGAAGGACGAAAGCTAGGG 687 Sbjct 654 ACACCGGTGGCGAAAGCGCTTTACTGGGCCAGACCTGACACTGAAGGACGAAAGCTAGGG 713 Query 688 GAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCCGTAAACGATGGAGACTAGGTGTAG 747 Sbjct 714 GAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCTGTAAACGATGGAGACTAGGTGTAG 773 Query 748 CACGTATCGACCCG 761 Sbjct 774 CACGTATCGACCCG 787 R Uncultured cyanobacterium clone UFDCBA_30f 16S ribosomal RNA gene, partial sequence Length=1454 Score = 1240 bits (671), Expect = 0.0 Identities = 721/745 (96%), Gaps = 3/745 (0%) Strand=Plus/Minus Query 28 CTA-GACTAC-GGGGTATCTAATCCCTTTCGCTCCCCTAGCTTTCGTCCTTCACTGTCAG 85 Sbjct 747 CTAGGACTACAGGGGTATCTAATCCCTTTCGCTCCCCTAGCTTTCGTCCTTCAGTGTCAG 688 Query 86 GGCTGGCCCAGTAAAGCGCTTTCGCCACCGGTGTTCTTCCCGATATCTACGCATTTCACC 145 Sbjct 687 GTCTGGCCCAGTAAAGCGCTTTCGCCACCGGTGTTCTTCCCGATATCTACGCATTTCACC 628 Query 146 GCTACACCGGGAATTCCCTTTACCCCTACCAGCCTCTAGCTAACCAGTTTCCACTGCCTA 205 Sbjct 627 GCTACACCGGGAATTCCCTTTACCCCTACCAACCTCTAGCTTGCCAGTTTCCACTGCCTT 568 Query 206 CCCACAGTTGAGCTGTGATCTTTAACAGCAGACTTAGTTCGCCACCTACAGACTCTTTAC 265 Sbjct 567 CCCACAGTTGAGCTGTGATCTTTAACAGCAGACTTAGCTGGCCACCTACAGACGCTTTAC 508 Query 266 GCCCAATTATTCCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCTGCTGGCACGGAGT 325 Sbjct 507 GCCCAATCATTCCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCTGCTGGCACGGAGT 448
86 64 Query 326 TAGCCGAGGCTTATTCGTCAAGTACCGTCAGAGCTTCTTCCTTGACAAAAGGGGTTTACG 385 Sbjct 447 TAGCCGAGGCTTATTCGTCAAGTACCGTCAGAGCTTCTTCCTTGACAAAAGGGGTTTACG 388 Query 386 ATCCAAAAACCTTCCTCCCCCACGCGGTCTTGCTCCGTCAGGCTTTCGCCCATTGCGGAA 445 Sbjct 387 ATCCAAAAACCTTCCTCCCCCACGCGGTCTTGCTCCGTCAGGCTTTCGCCCATTGCGGAA 328 Query 446 AATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCCAGTGTGGCTGC 505 Sbjct 327 AATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCCAGTGTGGCTGC 268 Query 506 TCATCCTCTCAGACCAGCTACTGATCGTCGCCTTGGTGCGCTTTTACCACACCAACTAGC 565 Sbjct 267 TCATCCTCTCAGACCAGCTACTGATCGTCGCCTTGGTACGCTTTTACCTTACCAACTAGC 208 Query 566 TAATCAGACGCGAGCTCATCTGTGGGCAATAAATCTTTTACCTTTCAGCTCATCCAGAAT 625 Sbjct 207 TAATCAGACGCGAGCTCATCTGTGGGCAATAAATCTTTTACCTTTCGGCTCATCCGGTAT 148 Query 626 TAGCCACCGTTTCCAGTGGTTATGCCCGACCCACAGGCAGATTCTCACGCGTTACTCACC 685 Sbjct 147 TAGCCACCGTTTCCAGTAGTTGTCCCCGACCCACAGGCAGATTCTCACGCGTTACTCACC 88 Query 686 CGTCCGCCACTCACTCCGAAGAGTTCGCTCGACTTGCATGTGTTAAGCAGACCGGCAGCG 745 Sbjct 87 CGTCCGCCACTCACTCCGAAGAGTTCGTTCGACTTGCATGTGTTAAGCAGACCGCCAGCG 28 Query 746 TTCATCCTGAGCCAGGAATCAAACT 770 Sbjct 27 TTCATCCTGAGCCAGGA-TCAAACT Uncultured cyanobacterium clone BB1S1... Uncultured cyanobacterium clone BB1S1(3) Pseudoscillatoria coralii BgP10 4S FJ... Uncultured cyanobacterium clone BB1S1(2) Uncultured cyanobacterium UFDCBA 30f... Uncultured cyanobacterium clone UNFR... Uncultured cyanobacterium clone UNFR... U Osc 27F-809R KK Lyngbya birgei CCC S ribosomal RNA gene, partial sequence Length=706 Score = 1249 bits (676), Expect = 0.0 Identities = 697/707 (98%), Gaps = 2/707 (0%) Strand=Plus/Plus Query 29 GTGTCTTCGGACACAGTGGCGGACGGGTGAGTAACGCGTGAGAATCTACCTTCAGGTCGG 88 Sbjct 1 GTGTCTTCGGACACAGTGGCGGACGGGTGAGTAACGCGTGAGAATCTACCTTCAGGTCGG 60 Query 89 GGATAACCCCTGGAAACGGGGGCTAATACCGGATAGGTCGAAAGATGAAAGGTTAACTGC 148 Sbjct 61 GGATAACCCCTGGAAACGGGGGCTAATACCGGATAGGTCGAAAGATGAAAGGTTAACTGC 120 Query 149 CTGAAGAGGAGCTCGCGTCTGATTAGCTAGTAGGTGTGGTAAGAGCGCACCTAGGCGACG 208 Sbjct 121 CTGAAAAGGGGCTCGCGTCTGATTAGCTAGTAGGTGTGGTAAGAGCGCACCTAGGCGACA 180 Query 209 ATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCCAGACTC 268 Sbjct 181 ATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCCAAACTC 240
87 65 Query 269 CTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACC 328 Sbjct 241 CTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACC 300 Query 329 GCGTGGGGGAGGAAGGCCTTTGGGTTGTAAACCCCTTTTGTTAGGGAAGaaaaaaaTGAC 388 Sbjct 301 GCGGGGGGGAGGAAGGCCTTTGGGTTGTAAACCCCTTTTGTTAGGGAAAAAAAAAATGAC 360 Query 389 GGTACCTAACGAATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAG 448 Sbjct 361 GGTACCTAACGAATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAG 420 Query 449 GCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGCCCGGCAAGTCTGCT 508 Sbjct 421 GCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGCCCGGCAAGTCTGCT 480 Query 509 GTCAAAGACCACAGCTCAACTGTGGAAAGGCAGTGGAAACTGCAAGGCTAGAGTGCGGTA 568 Sbjct 481 GTCAAAGACCACAGCTCAACTGTGGAAAGGCAGTGGAAACTGCAAGGCTAGAGTGCGGTA 540 Query 569 GGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGAGATTGGGAAGAACACCGGT 628 Sbjct 541 GGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGAGATTGGAAAGAACACCGGT 600 Query 629 GGCGAAAGCGCTCTACTGGGCCGCAACTGACACTGATGGACGAAAGCTAGGGGAGCGAAT 688 Sbjct 601 GGCGAAAGCGCTCTACTGGGCCGCAACTGACACTGATGGACGAAAGCTAGGGGAGCGAAT 660 Query 689 GGGATTAGATACCCC-AGTAGTCCTAGCTGTAAACGATGAGAAACTA 734 Sbjct 661 GGGATTAGATCCCCCCAGTAGTCCTAGCTGTAAACGATG-GAAACTA 706 R Lyngbya birgei CCC S ribosomal RNA gene, partial sequence Length=706 Score = 385 bits (208), Expect = 2e-103 Identities = 341/403 (84%), Gaps = 18/403 (4%) Strand=Plus/Plus Query 151 TCGCGT-TGATTACCTAGTAGGTGTGGTAAGAGCGCACCTTTGCGACTATCAGTTGCTGG 209 Sbjct 133 TCGCGTCTGATTAGCTAGTAGGTGTGGTAAGAGCGCACCTAGGCGACAATCAGTAGCTGG 192 Query 210 TCTGACAGGATGA-CTTGCCGGGCTGGGACTGAAACACTGCCCACACTCCTACGGGAGGC 268 Sbjct 193 TCTGAGAGGATGAGC-AGCCACACTGGGACTGAGACACGGCCCAAACTCCTACGGGAGGC 251 Query 269 AGCATTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGATCAACACCGCGTGGGGCAG 328 Sbjct 252 AGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACCGCGGGGGGGAG 311 Query 329 GAAGGCCTTTGGGTTTTTT-CTTCCTTTTGTTAGGGAAGaaaaaaaTGAAAGG-ACCTTA 386 Sbjct 312 GAAGGCCTTTGGGTTGTAAACC-CCTTTTGTTAGGGAAAAAAAAAATGAC-GGTACCTAA 369 Query 387 CTAATCACCCTCGGCTAACTCCGTGCCAGCATCCGCGTTAATACAGAAGACGCAAGCGCT 446 Sbjct 370 CGAATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTT 429 Query 447 ATCCGGAATTATGGGGCGGTAAAGGGTCCGTGGGTGGCTCTGCCTGCCTG-TCGTCTTAG 505 Sbjct 430 ATCCGGAATTATTGGGCG-TAAAGCGTCCGTAGGTGGCCCGGCAAGTCTGCT-GTCAAAG 487 Query 506 ACCA-AGCCTCGGAA-TGTGGAACGGGCAGTGGAGAA-TGCAA 545 Sbjct 488 ACCACAGC-TC--AACTGTGGAAAGG-CAGTGGA-AACTGCAA 525
88 Oscillatoria spongeliae 310P1 AY Oscillatoria spongeliae 310P1 AY Oscillatoria spongeliae 31P1 AY Oscillatoria spongeliae 32P1 AY Oscillatoria spongeliae 520bg AF Microcoleus chthonoplastes CCY0602 GQ... Oscillatoria sp. MPI 990BR03 AF KK 27F-809R Phormidium animale PMC PMC Phormidium animale NTMP03 NTMP03 GU Lyngbya birgei CCC 333 EU Arthrospira sp. CCC 538 EU Bal pse Uncultured cyanobacterium clone Mat-CYANO-S19 16S ribosomal RNA gene, partial sequence Length=1413 Score = 867 bits (469), Expect = 0.0 Identities = 501/516 (97%), Gaps = 4/516 (0%) Strand=Plus/Plus Query 36 AGTCGAACGGAGTGCTTTTCGGGCACTTAGTGGCGGACGGGTGAGTAACGCGTGAGAACC 95 Sbjct 33 AGTCGAACGGAGTGC-CTTCGGGCACTTAGTGGCGGACGGGTGAGTAACGCGTGAGGATC 91 Query 96 TG-CCTTCAGGTCGGGGATAACGGCTGGAAACGGTCGCTAATACCGGATGTGCCGAGAGG 154 Sbjct 92 TGCCCTT-AGGTCGGGGATAACGACTGGAAACGGTCGCTAATACCGGATATGCCGAGAGG 150 Query 155 TGAAAGGTTAACTGCCTGAGGATGAGCTCGCGTCGGATTAGCTAGTTGGTGGGGTAAGGG 214 Sbjct 151 TGAAAGGTTAACTGCCTGAGGATGAGCTCGCGTCGGATTAGCTAGTTGGTGGGGTAAGGG 210 Query 215 CCTACCAAGGCGACGATCCGTAGCTGGTTTGAGAGGATGACCAGCCACACTGGGACTGAG 274 Sbjct 211 CCTACCAAGGCGACGATCCGTAGCTGGTTTGAGAGGATGACCAGCCACACTGGGACTGAG 270 Query 275 ACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGCAAGCC 334 Sbjct 271 ACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGCAAGCC 330 Query 335 TGACGGAGCAACGCCGCGTGCGGGAGGAAGGCCTGTGGGTTGTAAACCGCTTTTCTCAAG 394 Sbjct 331 TGACGGAGCAACGCCGCGTGGGGGAGGAAGGCCTGTGGGTTGTAAACCTCTTTTCTCAAG 390 Query 395 GAAGAAGTCAGTGACGGTACTTGAGGAATCAGCATCGGCTAACTCCGTGCCAGCAGCCGC 454 Sbjct 391 GAAGAAGCCAGTGACGGTACTTGAGGAATCAGCATCGGCTAACTCCGTGCCAGCAGCCGC 450 Query 455 GGTAAGACGGAGGATGCAAGCGTTATCCGGAATGATTGGGCGTAAAGCGTCCGCAGGTGG 514 Sbjct 451 GGTAAGACGGAGGATGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGCAGGTGG 510 Query 515 TTATACAAGTCTGCCGTTAAAGCGAGGGGCTTAACT 550 Sbjct 511 TTATACCAGTCTGCCGTTAAAGCGTGGG-CTTAACT 545 R Uncultured cyanobacterium clone Mat-CYANO-S19 16S ribosomal RNA
89 67 gene, partial sequence Length=1413 Score = 1245 bits (674), Expect = 0.0 Identities = 725/748 (96%), Gaps = 10/748 (1%) Strand=Plus/Minus Query 16 CATCGTTTACGGCTAGGACTACAGGGGGTATCTAATCCCTTTCGCTCCCCTAGCTTTCGT 75 Sbjct 740 CATCGTTTACGGCTAGGACTACA-GGGGTATCTAATCCCTTTCGCTCCCCTAGCTTTCGT 682 Query 76 CCCTCATTAGAT-AGACAAGGCCCAGTAGAGCGCTTTCGCCACTGGTGTTCTTCCCGATA 134 Sbjct 681 CCCTCAGT-G-TCAGTCAAGGCCCAGTAGAGCGCTTTCGCCACTGGTGTTCTTCCCGATA 624 Query 135 TCTACGCATTTCACCGCTACACCGGGAATTCCCTCTACCCCTACCTCACTCTAGTCATAC 194 Sbjct 623 TCTACGCATTTCACTGCTACACCGGG-ATTCCCTCTACCCCTACCTCACTCTAGTCATAC 565 Query 195 AGTTTCCACCGCCCTTATGGAGTTAAGCCCCACGCTTTAACGGCAGACTTGTATAACCAC 254 Sbjct 564 AGTTT-CACCGCCTTTATGGAGTTAAG-CCCACGCTTTAACGGCAGACTGGTATAACCAC 507 Query 255 CTGCGGACGCTTTACGCCCAATCATTCCGGATAACGCTTGCATCCTCCGTCTTACCGCGG 314 Sbjct 506 CTGCGGACGCTTTACGCCCAATAATTCCGGATAACGCTTGCATCCTCCGTCTTACCGCGG 447 Query 315 CTGCTGGCACGGAGTTAGCCGATGCTGATTCCTCAAGTACCGTCACTGACTTCTTCCTTG 374 Sbjct 446 CTGCTGGCACGGAGTTAGCCGATGCTGATTCCTCAAGTACCGTCACTGGCTTCTTCCTTG 387 Query 375 AGAAAAGCGGTTTACAACCCACAGGCCTTCCTCCCGCACGCGGCGTTGCTCCGTCAGGCT 434 Sbjct 386 AGAAAAGAGGTTTACAACCCACAGGCCTTCCTCCCCCACGCGGCGTTGCTCCGTCAGGCT 327 Query 435 TGCGCCCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCA 494 Sbjct 326 TGCGCCCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCA 267 Query 495 GTCCCAGTGTGGCTGGTCATCCTCTCAAACCAGCTACGGATCGTCGCCTTGGTAGGCCCT 554 Sbjct 266 GTCCCAGTGTGGCTGGTCATCCTCTCAAACCAGCTACGGATCGTCGCCTTGGTAGGCCCT 207 Query 555 TACCCCACCAACTAGCTAATCCGACGCGAGCTCATCCTCAGGCAGTTAACCTTTCACCTC 614 Sbjct 206 TACCCCACCAACTAGCTAATCCGACGCGAGCTCATCCTCAGGCAGTTAACCTTTCACCTC 147 Query 615 TCGGCACATCCGGTATTAGCGACCGTTTCCAGCCGTTATCCCCGACCTGAAGG-CAGATT 673 Sbjct 146 TCGGCATATCCGGTATTAGCGACCGTTTCCAGTCGTTATCCCCGACCT-AAGGGCAGATC 88 Query 674 CTCACGCGTTACTCACCCGTCCGCCACTAAGTGCCCGAAAGCACTCCGTTCGACTTGCAT 733 Sbjct 87 CTCACGCGTTACTCACCCGTCCGCCACTAAGTGCCCGAAGGCACTCCGTTCGACTTGCAT 28 Query 734 GTGTTAAGCACGCCGCCAGGCGTTCATC 761 Sbjct 27 GTGTTAAGCACGCCGCCAG-CGTTCATC 1
90 Limnothrix redekei 2LT25S01 FM Limnothrix sp. CENA109 EF Limnothrix sp. CENA110 EF Limnothrix sp. CENA111 EF Limnothrix redekei 165c AJ Bal pse 27F-809R Uncultured cyanobacterium clone Mat-C... Filamentous thermophilic cyanobacteri... Uncultured cyanobacterium clone BAC T... Leptolyngbya sp. 2LT21S03 FM ZD Geitlerinema sp. CR_13M 16S ribosomal RNA gene, partial sequence Length=1181 Score = 1127 bits (610), Expect = 0.0 Identities = 655/677 (96%), Gaps = 3/677 (0%) Strand=Plus/Plus Query 82 CAGGAGGGGGACAACCACTGGAAACGGTGGCTAATACCCCATAAGCCGAGAGGTAAAAAG 141 Sbjct 1 CAGGAGGGGGACAACCATTGGAAACGGTGGCTAATACCCCATAAGCCGAGAGGTGAAAAG 60 Query 142 GAATCCGCCTGTGGAGGAGCTCGCGTCTGATTAGCTAGTAGGTGAGGTAAGAGCTTACCT 201 Sbjct 61 GAATCCGCCTGTGGAGGAGCTCGCGTCTGATTAGCTAGTAGGTAAGGTAAAAGCTTACCT 120 Query 202 AGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGC 261 Sbjct 121 AGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGC 180 Query 262 CCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGA 321 Sbjct 181 CCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGA 240 Query 322 GCCAGACCGCGTGAGGGAAGAAGGCCTTTGGGTTGTAAACCTCTTTTCTCAGGGAAGAAG 381 Sbjct 241 GCCAGACCGCGTGAGGGAAGAAGGCCCTTGGGTTGTAAACCTCTTTTCTCAAGGAAGAAG 300 Query 382 AAAGTGACGGTACCTGAGG-AATCAG-CCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAA 439 Sbjct 301 CAAGTGACGGTACTTGAGGCAAT-AGGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAA 359 Query 440 GACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGTGATT 499 Sbjct 360 GACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGTGACT 419 Query 500 CAAGTCTGCTGTCAAAACCCAGGGCTTAACTCTGGATCGGCAGTGGAAACTGAATGACTA 559 Sbjct 420 CAAGTCTGCTGTCAAAAACTAGGGCTTAACTCTAGAGAGGCAGTGGAAACTGAGTAACTA 479 Query 560 GAGTGCGGCAGGGGCAGAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGAGATCGGGAA 619 Sbjct 480 GAGAGCGGCAGGGGCAGAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGRGATCGGGAA 539 Query 620 GAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGCGACTGACACTCACAGGACGAAAGCTA 679 Sbjct 540 GAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGCGACTGACACTCACAGGACGAAAGCTA 599 Query 680 GGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCTGTAAACGATGGAAACTAGGTG 739 Sbjct 600 GGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCCGTAAACGATGGAAACTAGGTG 659
91 69 Query 740 TGGCCTGTATCGACCCG 756 Sbjct 660 TGGCCTGTATCGACCCG 676 R Geitlerinema sp. CR_13M 16S ribosomal RNA gene, partial sequence Length=1181 Score = 1064 bits (576), Expect = 0.0 Identities = 624/647 (96%), Gaps = 5/647 (0%) Strand=Plus/Minus Query 11 TCGTTT-C-GCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCCT 68 Sbjct 646 TCGTTTACGGCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCCT 587 Query 69 GTGAGTGTCAGTCGCGGCCCAGCAGAGCGCTTTCGCCACCGGTGTTCTTCCCGATCTCTA 128 Sbjct 586 GTGAGTGTCAGTCGCGGCCCAGCAGAGCGCTTTCGCCACCGGTGTTCTTCCCGATCYCTA 527 Query 129 CGCATTTCACCGCTACACCGGGAATTCCCTCTGCCCCTGCCGCACTCTAGTCATTCAGTT 188 Sbjct 526 CGCATTTCACCGCTACACCGGGAATTCCCTCTGCCCCTGCCGCTCTCTAGTTACTCAGTT 467 Query 189 TCCACTGCCGATCCAGAGTTAAGCCCTGGGTTTTGACAGCAGACTTGAATCACCACCTAC 248 Sbjct 466 TCCACTGCCTCTCTAGAGTTAAGCCCTAGTTTTTGACAGCAGACTTGAGTCACCACCTAC 407 Query 249 GGACGCTTTACGCCCAATAATTCCGGATAACGCTTGCCTCCTCCGTCTTACCGCGGCTGC 308 Sbjct 406 GGACGCTTTACGCCCAATAATTCCGGATAACGCTTGCCTCCTCCGTCTTACCGCGGCTGC 347 Query 309 TGGCACGGAGTTAGCCGAGG-CTGATT-CCTCAGGTACCGTCACTTTCTTCTTCCCTGAG 366 Sbjct 346 TGGCACGGAGTTAGCCGAGGCCT-ATTGCCTCAAGTACCGTCACTTGCTTCTTCCTTGAG 288 Query 367 AAAAGAGGTTTACAACCCAAAGGCCTTCTTCCCTCACGCGGTCTGGCTCCGTCAGGCTTT 426 Sbjct 287 AAAAGAGGTTTACAACCCAAGGGCCTTCTTCCCTCACGCGGTCTGGCTCCGTCAGGCTTT 228 Query 427 CGCCCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGT 486 Sbjct 227 CGCCCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGT 168 Query 487 CCCAGTGTGGCTGCTCATCCTCTCAGACCAGCTACTGATCGTCGCCTAGGTAAGCTCTTA 546 Sbjct 167 CCCAGTGTGGCTGCTCATCCTCTCAGACCAGCTACTGATCGTCGCCTAGGTAAGCTTTTA 108 Query 547 CCTCACCTACTAGCTAATCAGACGCGAGCTCCTCCACAGGCGGATTCCTTTTTACCTCTC 606 Sbjct 107 CCTTACCTACTAGCTAATCAGACGCGAGCTCCTCCACAGGCGGATTCCTTTTCACCTCTC 48 Query 607 GGCTTATGGGGTATTAGCCACCGTTTCCAGTGGTTGTCCCCCTCCTG 653 Sbjct 47 GGCTTATGGGGTATTAGCCACCGTTTCCAATGGTTGTCCCCCTCCTG 1 Şif Geitlerinema sp. PCC 7105 small subunit ribosomal RNA gene, partial sequence Length=1409 Score = 1347 bits (729), Expect = 0.0 Identities = 737/740 (99%), Gaps = 3/740 (0%) Strand=Plus/Plus Query 18 CATGC-AGTCG-ACG-AGTCTTCGGACTTAGTGGCGGACGGGTGAGTAACGCGTGAGAAC 74
92 70 Sbjct 27 CATGCAAGTCGAACGAAGTCTTCGGACTTAGTGGCGGACGGGTGAGTAACGCGTGAGAAC 86 Query 75 CTGCCTCGAGGAGGGGGATAACAGCGGGAAACTGCTGCTAATACCCCATATGCCGAAAGG 134 Sbjct 87 CTGCCTCGAGGAGGGGGATAACAGCGGGAAACTGCTGCTAATACCCCATATGCCGAAAGG 146 Query 135 TGAAAAGAAATTTGCCTTGAGAGGGGCTCGCGTCCGATTAGCTAGTTGGTGAGGTAAGAG 194 Sbjct 147 TGAAAAGAAATTTGCCTTGAGAGGGGCTCGCGTCCGATTAGCTAGTTGGTGAGGTAAGAG 206 Query 195 CTTACCAAGGCGACGATCGGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAG 254 Sbjct 207 CTTACCAAGGCGACGATCGGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAG 266 Query 255 ACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCC 314 Sbjct 267 ACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCC 326 Query 315 TGACGGAGCAACGCCGCGTGGGGGAAGAAGGCCTTTGGGTTGTAAACTCCTTTTCTCAGG 374 Sbjct 327 TGACGGAGCAACGCCGCGTGGGGGAAGAAGGCCTTTGGGTTGTAAACTCCTTTTCTCAGG 386 Query 375 GAAGAAGAACTGACGGTACCTGAGGAATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCG 434 Sbjct 387 GAAGAAGAACTGACGGTACCTGAGGAATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCG 446 Query 435 GTAATACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTTCGTAGGCGGC 494 Sbjct 447 GTAATACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTTCGTAGGCGGC 506 Query 495 GTTTCAAGTCTGCTGTCAAAGGCCGAGGCTCAACTTCGGAAAGGCAGTGGAAACTGAAAA 554 Sbjct 507 GTTTCAAGTCTGCTGTCAAAGGCCGAGGCTCAACTTCGGAAAGGCAGTGGAAACTGAAAA 566 Query 555 GCTAGAGGTCGGTAGGGGCAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTG 614 Sbjct 567 GCTAGAGGTCGGTAGGGGCAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTG 626 Query 615 GGAAGAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGAACCTGACGCTGAGGGACGAAAG 674 Sbjct 627 GGAAGAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGAACCTGACGCTGAGGGACGAAAG 686 Query 675 CTAGGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCCGTAAACGATGGAGACTAG 734 Sbjct 687 CTAGGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCCGTAAACGATGGAGACTAG 746 Query 735 GTGTTGCCTGTATCGACCCG 754 Sbjct 747 GTGTTGCCTGTATCGACCCG 766 R Geitlerinema sp. PCC 7105 small subunit ribosomal RNA gene, partial sequence Length=1409 Score = 1360 bits (736), Expect = 0.0 Identities = 736/736 (100%), Gaps = 0/736 (0%) Strand=Plus/Minus Query 15 TCGTTTACGGCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCCC 74 Sbjct 736 TCGTTTACGGCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCCC 677 Query 75 TCAGCGTCAGGTTCGGCCCAGCAGAGCGCTTTCGCCACCGGTGTTCTTCCCAATATCTAC 134 Sbjct 676 TCAGCGTCAGGTTCGGCCCAGCAGAGCGCTTTCGCCACCGGTGTTCTTCCCAATATCTAC 617 Query 135 GCATTTCACCGCTACACTGGGAATTCCCTCTGCCCCTACCGACCTCTAGCTTTTCAGTTT 194 Sbjct 616 GCATTTCACCGCTACACTGGGAATTCCCTCTGCCCCTACCGACCTCTAGCTTTTCAGTTT 557 Query 195 CCACTGCCTTTCCGAAGTTGAGCCTCGGCCTTTGACAGCAGACTTGAAACGCCGCCTACG 254
93 71 Sbjct 556 CCACTGCCTTTCCGAAGTTGAGCCTCGGCCTTTGACAGCAGACTTGAAACGCCGCCTACG 497 Query 255 AACGCTTTACGCCCAATAATTCCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCTGCT 314 Sbjct 496 AACGCTTTACGCCCAATAATTCCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCTGCT 437 Query 315 GGCACGGAGTTAGCCGAGGCTGATTCCTCAGGTACCGTCAGTTCTTCTTCCCTGAGAAAA 374 Sbjct 436 GGCACGGAGTTAGCCGAGGCTGATTCCTCAGGTACCGTCAGTTCTTCTTCCCTGAGAAAA 377 Query 375 GGAGTTTACAACCCAAAGGCCTTCTTCCCCCACGCGGCGTTGCTCCGTCAGGCTTTCGCC 434 Sbjct 376 GGAGTTTACAACCCAAAGGCCTTCTTCCCCCACGCGGCGTTGCTCCGTCAGGCTTTCGCC 317 Query 435 CATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCCA 494 Sbjct 316 CATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCCA 257 Query 495 GTGTGGCTGCTCATCCTCTCAGACCAGCTACCGATCGTCGCCTTGGTAAGCTCTTACCTC 554 Sbjct 256 GTGTGGCTGCTCATCCTCTCAGACCAGCTACCGATCGTCGCCTTGGTAAGCTCTTACCTC 197 Query 555 ACCAACTAGCTAATCGGACGCGAGCCCCTCTCAAGGCAAATTTCTTTTCACCTTTCGGCA 614 Sbjct 196 ACCAACTAGCTAATCGGACGCGAGCCCCTCTCAAGGCAAATTTCTTTTCACCTTTCGGCA 137 Query 615 TATGGGGTATTAGCAGCAGTTTCCCGCTGTTATCCCCCTCCTCGAGGCAGGTTCTCACGC 674 Sbjct 136 TATGGGGTATTAGCAGCAGTTTCCCGCTGTTATCCCCCTCCTCGAGGCAGGTTCTCACGC 77 Query 675 GTTACTCACCCGTCCGCCACTAAGTCCGAAGACTTCGTTCGACTTGCATGTGTTAGGCAC 734 Sbjct 76 GTTACTCACCCGTCCGCCACTAAGTCCGAAGACTTCGTTCGACTTGCATGTGTTAGGCAC 17 Query 735 GCCGCCAGCGTTCATC 750 Sbjct 16 GCCGCCAGCGTTCATC 1 Geitlerinema sp. PCC 7105 AB Phormidium lucidum BDU GU Geitlerinema sp. PCC 7105 AF Sif 27F-809R Jaaginema pseudogeminatum NTMP02 GU81... Geitlerinema sp. Flo1 FJ Geitlerinema sp. MBIC10006 AB Geitlerinema sp. A28DM FJ Geitlerinema sp. BBD HS217 EF Geitlerinema sp. W-1 EF Geitlerinema sp. BBD P2b-1 EF Geitlerinema sp. BBD DQ Bal ip Geitlerinema sp. PCC S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 and 23S rrna gene (partial), strain PCC 8501 Length=2024
94 72 Score = 1343 bits (727), Expect = 0.0 Identities = 740/746 (99%), Gaps = 2/746 (0%) Strand=Plus/Plus Query 16 TACCATGC-AGTCG-ACGGAGTCCTTCGGGACTTAGTGGCGGACGGGTGAGTAACGCGTG 73 Sbjct 4 TACCATGCAAGTCGAACGGAGTGCTTCGGCACTTAGTGGCGGACGGGTGAGTAACGCGTG 63 Query 74 AGAATCTGCCCTTAGGACGGGGACAACAGCTGGAAACGGCTGCTAATACCCGATGTGCCG 133 Sbjct 64 AGAATCTGCCCTTAGGACGGGGACAACAGCTGGAAACGGCTGCTAATACCCGATGTGCCG 123 Query 134 AGAGGTGAAACATTTTATGGCCTGAGGATGAGCTCGCGTCTGATTAGCTAGTTGGTGGGG 193 Sbjct 124 AGAGGTGAAACATTTTATGGCCTGAGGATGAGCTCGCGTCTGATTAGCTAGTTGGTGGGG 183 Query 194 TAAGGGCCTACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGATCAGCCACACTGGG 253 Sbjct 184 TAAGAGCCTACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGATCAGCCACACTGGG 243 Query 254 ACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCG 313 Sbjct 244 ACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCG 303 Query 314 CAAGCCTGACGGAGCAAGACCGCGTGCGGGAGGACGGCCTATTGGTTGTAAACCGCTTTT 373 Sbjct 304 CAAGCCTGACGGAGCAAGACCGCGTGCGGGAGGACGGCCTATTGGTTGTAAACCGCTTTT 363 Query 374 GATAGGGAAGAAGTTCTGACGGTACCTATCGAATCAGCATCGGCTAACTCCGTGCCAGCA 433 Sbjct 364 GATAGGGAAGAAGTACTGACGGTACCTATCGAATCAGCATCGGCTAACTCCGTGCCAGCA 423 Query 434 GCCGCGGTAATACGGAGGATGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTA 493 Sbjct 424 GCCGCGGTAATACGGAGGATGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTA 483 Query 494 GGTGGTTGTTCAAGTCTGCTGTTAAAGCGTGCGGCTTAACCGCATACCAGCAGTGGAAAC 553 Sbjct 484 GGTGGTTGTTCAAGTCTGCTGTTAAAGCGTGCGGCTTAACCGCATACCAGCAGTGGAAAC 543 Query 554 TGGGCGACTAGAGTGCGGTAGGGGTCAGGGGAATTCCCAGTGTAGCGGTGAAATGCGTAG 613 Sbjct 544 TGGGCGACTAGAGTGCGGTAGGGGTCAGGGGAATTCCCAGTGTAGCGGTGAAATGCGTAG 603 Query 614 ATATTGGGAAGAACACCGGTGGCGAAAGCGCCTGACTGGACCTGCACTGACACTGAGGGA 673 Sbjct 604 ATATTGGGAAGAACACCGGTGGCGAAAGCGCCTGACTGGACCTGCACTGACACTGAGGGA 663 Query 674 CGAAAGCTAGGGGAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCCGTAAACGATGGA 733 Sbjct 664 CGAAAGCTAGGGGAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCCGTAAACGATGGA 723 Query 734 CACTAGGTGTTGCCCGTATCGACCCG 759 Sbjct 724 CACTAGGTGTTGCCCGTATCGACCCG 749 R Geitlerinema sp. PCC S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 and 23S rrna gene (partial), strain PCC 8501 Length=2024 Score = 1293 bits (700), Expect = 0.0 Identities = 716/723 (99%), Gaps = 3/723 (0%) Strand=Plus/Minus Query 12 CATCGTTTACGGCTAGGACTACA-GGGTATCTAATCCCTTTCGCTCCCCTAGCTTTCGTC 70 Sbjct 721 CATCGTTTACGGCTAGGACTACAGGGGTATCTAATCCCTTTCGCTCCCCTAGCTTTCGTC 662 Query 71 CCTCAGTGTCAGTGCAGGTCCAGTCAGGCGCTTTCGCCACCGGTGTTCTTCCCAATATCT 130
95 73 Sbjct 661 CCTCAGTGTCAGTGCAGGTCCAGTCAGGCGCTTTCGCCACCGGTGTTCTTCCCAATATCT 602 Query 131 ACGCATTTCACCGCTACACTGGGAATTCCCCTGACCCCTACCGCACTCTAGTCGCCCAGT 190 Sbjct 601 ACGCATTTCACCGCTACACTGGGAATTCCCCTGACCCCTACCGCACTCTAGTCGCCCAGT 542 Query 191 TTCCACTGCTGGTATGCGGTTAAGCCGCACGCTTTAACAGCAGACTTGAACAACCACCTA 250 Sbjct 541 TTCCACTGCTGGTATGCGGTTAAGCCGCACGCTTTAACAGCAGACTTGAACAACCACCTA 482 Query 251 CGGACGCTTTACGCCCAATAATTCCGGATAACGCTTGCATCCTCCGTATTACCGCGGCTG 310 Sbjct 481 CGGACGCTTTACGCCCAATAATTCCGGATAACGCTTGCATCCTCCGTATTACCGCGGCTG 422 Query 311 CTGGCACGGAGTTAGCCGATGCTGATTCGATAGGTACCGTCAGAACTTCTTCCCTATCAA 370 Sbjct 421 CTGGCACGGAGTTAGCCGATGCTGATTCGATAGGTACCGTCAGTACTTCTTCCCTATCAA 362 Query 371 AAGCGGTTTACAACCAATAGGCCGTCCTCCCGCACGCGGTCTTGCTCCGTCAGGCTTGCG 430 Sbjct 361 AAGCGGTTTACAACCAATAGGCCGTCCTCCCGCACGCGGTCTTGCTCCGTCAGGCTTGCG 302 Query 431 CCCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCC 490 Sbjct 301 CCCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCC 242 Query 491 CAGTGTGGCTGATCATCCTCTCAGACCAGCTACTGATCGTCGCCTTGGTAGGCCCTTACC 550 Sbjct 241 CAGTGTGGCTGATCATCCTCTCAGACCAGCTACTGATCGTCGCCTTGGTAGGCTCTTACC 182 Query 551 CCACCAACTAGCTAATCAGACGCGAGCTCATCCTCAGGCCATAAAATGTTTCACCTCTCG 610 Sbjct 181 CCACCAACTAGCTAATCAGACGCGAGCTCATCCTCAGGCCATAAAATGTTTCACCTCTCG 122 Query 611 GCACATCGGGTATTAGCAGCCGTTTCCAGCTGTTGTCCCCGTCCTAAGGGCAGATTCTCA 670 Sbjct 121 GCACATCGGGTATTAGCAGCCGTTTCCAGCTGTTGTCCCCGTCCTAAGGGCAGATTCTCA 62 Query 671 CGCGTTACTCACCCGTCCGCCACTAAGTCCCGAAGGACTCCGTTCGACTTGCATGTGTTA 730 Sbjct 61 CGCGTTACTCACCCGTCCGCCACTAAGTGCCGAAGCACTCCGTTCGACTTGCATG-GT-A 4 Query 731 AGC 733 Sbjct 3 AGC 1 47 Bal ip 27F-809R 44 Filamentous thermophilic cyanobacteri Filamentous thermophilic cyanobacteri(2) 80 Filamentous thermophilic cyanobacteri(3) Geitlerinema sp. PCC 8501 FM Cyanobacterium sp. (OS type I) L Oscillatoria sp. J-24-Osc AF Leptolynbya laminosa ETS-08 FM Leptolyngbya sp. MMG-1 AF KY Phormidium animale PMC partial 16S rrna gene, strain PMC Length=614 Score = 1129 bits (611), Expect = 0.0
96 74 Identities = 613/614 (99%), Gaps = 0/614 (0%) Strand=Plus/Plus Query 51 AGTGGCGGACGGGTGAGTAACGCGTGAGAATCTACCTTCAGGTCGGGGATAACCCCTGGA 110 Sbjct 1 AGTGGCGGACGGGTGAGTAACGCGTGAGAATCTACCTTCAGGTCGGGGATAACCCCTGGA 60 Query 111 AACGGGGGCTAATACCGGATAGGTCGAAAGATGAAAGGTTAACTGCCTGAAGAGGAGCTC 170 Sbjct 61 AACGGGGGCTAATACCGGATAGGTCGAAAGATGAAAGGTTAACTGCCTGAAGAGGGGCTC 120 Query 171 GCGTCTGATTAGCTAGTAGGTGTGGTAAGAGCGCACCTAGGCGACGATCAGTAGCTGGTC 230 Sbjct 121 GCGTCTGATTAGCTAGTAGGTGTGGTAAGAGCGCACCTAGGCGACGATCAGTAGCTGGTC 180 Query 231 TGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGC 290 Sbjct 181 TGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGC 240 Query 291 AGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACCGCGTGGGGGAGGAA 350 Sbjct 241 AGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACCGCGTGGGGGAGGAA 300 Query 351 GGCCTTTGGGTTGTAAACCCCTTTTGTTAGGGAAGaaaaaaaTGACGGTACCTAACGAAT 410 Sbjct 301 GGCCTTTGGGTTGTAAACCCCTTTTGTTAGGGAAGAAAAAAATGACGGTACCTAACGAAT 360 Query 411 CAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTTATCCG 470 Sbjct 361 CAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTTATCCG 420 Query 471 GAATTATTGGGCGTAAAGCGTCCGTAGGTGGCCCGGCAAGTCTGCTGTCAAAGACCACAG 530 Sbjct 421 GAATTATTGGGCGTAAAGCGTCCGTAGGTGGCCCGGCAAGTCTGCTGTCAAAGACCACAG 480 Query 531 CTCAACTGTGGAAAGGCAGTGGAAACTGCAAGGCTAGAGTGCGGTAGGGGTAGAGGGAAT 590 Sbjct 481 CTCAACTGTGGAAAGGCAGTGGAAACTGCAAGGCTAGAGTGCGGTAGGGGTAGAGGGAAT 540 Query 591 TCCCAGTGTAGCGGTGAAATGCGTAGAGATTGGGAAGAACACCGGTGGCGAAAGCGCTCT 650 Sbjct 541 TCCCAGTGTAGCGGTGAAATGCGTAGAGATTGGGAAGAACACCGGTGGCGAAAGCGCTCT 600 Query 651 ACTGGGCCGCAACT 664 Sbjct 601 ACTGGGCCGCAACT 614 R Phormidium animale PMC partial 16S rrna gene, strain PMC Length=614 Score = 1055 bits (571), Expect = 0.0 Identities = 591/601 (98%), Gaps = 0/601 (0%) Strand=Plus/Minus Query 91 TAGAGCGGTTTCGCCACCGGTGTTCTTCCCAATCTCTACGCATTTCACCGCTACACTGGG 150 Sbjct 601 TAGAGCGCTTTCGCCACCGGTGTTCTTCCCAATCTCTACGCATTTCACCGCTACACTGGG 542 Query 151 AATTCCCTCTACCCCTACCGCACTCTAGCCTTGCAGTTTCCACTGCCTTTCCACAGTTGA 210 Sbjct 541 AATTCCCTCTACCCCTACCGCACTCTAGCCTTGCAGTTTCCACTGCCTTTCCACAGTTGA 482 Query 211 GCTGTGGTCTTTGACAGCAGACTTGCCGGGCCACCTACGGACGCTTTACGCCCAATAATT 270 Sbjct 481 GCTGTGGTCTTTGACAGCAGACTTGCCGGGCCACCTACGGACGCTTTACGCCCAATAATT 422 Query 271 CCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCTGCTGGCACGGAATTAGCCGAGGCT 330 Sbjct 421 CCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCTGCTGGCACGGAGTTAGCCGAGGCT 362 Query 331 GATTCATTAGGGACCGTCAtttttttCTTCCCTAACAAAAGGGGTTTACAACCCAAAGGC 390 Sbjct 361 GATTCGTTAGGTACCGTCATTTTTTTCTTCCCTAACAAAAGGGGTTTACAACCCAAAGGC 302
97 75 Query 391 CTTCCTCCCCCACGCGGTCTTGCTCCGTCAGGCTTTCGCCCATTGCGGAAAATTCCCCAC 450 Sbjct 301 CTTCCTCCCCCACGCGGTCTTGCTCCGTCAGGCTTTCGCCCATTGCGGAAAATTCCCCAC 242 Query 451 TGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCCAGTGTGGCTGCTCATCCTCTC 510 Sbjct 241 TGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCCAGTGTGGCTGCTCATCCTCTC 182 Query 511 AGACCAGCTACTGATCGTCGCCTAGGTGCGCTCTTACCACACCTACTAGCTAATCAGACG 570 Sbjct 181 AGACCAGCTACTGATCGTCGCCTAGGTGCGCTCTTACCACACCTACTAGCTAATCAGACG 122 Query 571 CGAGCTCCTCTTCAGGGAGTTAACCTTTCATCTTTCGACCTATCCGGAATTAGCCCCCGA 630 Sbjct 121 CGAGCCCCTCTTCAGGCAGTTAACCTTTCATCTTTCGACCTATCCGGTATTAGCCCCCGT 62 Query 631 TTCCAGGGGTTAGCCCCGACCTGAAGGTAGATTCTCACGCGTTACTCAACCGTCCGCCAC 690 Sbjct 61 TTCCAGGGGTTATCCCCGACCTGAAGGTAGATTCTCACGCGTTACTCACCCGTCCGCCAC 2 Query 691 T 691 Sbjct 1 T Lyngbya birgei CCC 333 EU Arthrospira sp. CCC 538 EU Phormidium animale NTMP03 GU Phormidium animale PMC AJ KY 27F-809R Microcoleus chthonoplastes CCY0602 G... Oscillatoria sp. MPI 990BR03 MPI AF28... Oscillatoria spongeliae 310P1 AY Oscillatoria sp. Ck2 GQ Oscillatoria spongeliae SI04-46 EF Oscillatoria spongeliae SI04-40 EF U sal Halospirulina sp.'ccc Baja-95 Cl.3', 16S rrna gene Length=1444 Score = 1162 bits (629), Expect = 0.0 Identities = 714/753 (94%), Gaps = 16/753 (2%) Strand=Plus/Plus Query 16 TGC-AGTCG-ACGG-TCTCTTAGGAGAT-AGTGGCGGACGGGTGAGTAACGCGTGAGAAC 71 Sbjct 33 TGCAAGTCGAACGGAT-TCTTAGGA-ATCAGTGGCGGACGGGTGAGTAACACGTGAAAAC 90 Query 72 CTGCCTCTAGGACGGGGACAACAGAGGGAAACTTCTGCTAATTCCCGAT-ACGCCTTTGG 130 Sbjct 91 CTGCCTCTAGGACGGGGATAACAGAGGGAAACTTCTGCTAATTCCCGATGA-GCCTGCGG 149 Query 131 GTAAAA-GATTTATCGCCTAGAGAGGGGCTCGCGTCCGATTAGCTAGTTGGTGGGGTAAA 189 Sbjct 150 GTAAAAGGATTTATCGCCTAGAGAGGGGTTCGCGTCCGATTAGCTAGTTGGTGGGGT-AA 208 Query 190 AGCCT-ACCAAGGCAGCGATCGGTAGCTGTTCTGAGAGGAAGAACAGCCACACTGGGACT 248 Sbjct 209 AGGCTCACCAAGGCTGCGATCGGTAGCTGTTCTGAKAGGAAGAACAGCCACACTGGGACT 268
98 76 Query 249 GAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGCAA 308 Sbjct 269 GAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAA 328 Query 309 GCCTGACGGAGCAATACCGCGTGAGGGAAGAAGGTCTTTGGATCGTAAACCTCTTTTCTC 368 Sbjct 329 GCCTGACGGAGCAATACCGCGTGAGGGAAGAAGGTCTTTGGATCGTAAACCTCTTTTCTC 388 Query 369 AGGGAAGAAGATCTGACGGTACCTGAGGAATAAGCCTCGGCTAACTCCGTGCCAGCAGCC 428 Sbjct 389 AGGGAAGAAGTTCTGACGGTACCTGAGGAATAAGCCTCGGCTAACTCCGTGCCAGCAGCC 448 Query 429 GCGGTAATACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTTCGCAGGT 488 Sbjct 449 GCGGTAATACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTTCGCAGGT 508 Query 489 GGCTATTCAAGTCTGCTGTCAAAGACCAGAGCTCAACTCTGGGTAGGCGGTGGAAACTGA 548 Sbjct 509 GGCTCGTCAAGTCTGCTGTCAAAGACCGGAGCTTAACTCCGGGTAGGCGGTGGAAACTGA 568 Query 549 ATAGCTAGAGTGCGGTAGGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGAGA 608 Sbjct 569 AGAGCTAGAGTGCGGTAGGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGAGA 628 Query 609 TTGGGAAGAACATCGGTGGCGAAAGCGCTCTACT-GGACCGCAACTGACACTCAGGGACG 667 Sbjct 629 TTGGGAAGAACACCGGTGGCGAAAGCGCTCTACTAGG-CCGCAACTGACACTCAGGGACG 687 Query 668 AAAGCTAGGGGAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCCGTAAACGATGGAGA 727 Sbjct 688 AAAGCTAGGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCTGTAAACGATGG-GT 746 Query 728 AACTAGGCGTAGATTGTATCGACCCGAGC-GTG 759 Sbjct 747 A-CTAGGCGTAGCTTGTATCGACCCGAGCTGTG 778 R Halospirulina sp.'ccc Baja-95 Cl.3', 16S rrna gene Length=1444 Score = 1149 bits (622), Expect = 0.0 Identities = 709/749 (94%), Gaps = 16/749 (2%) Strand=Plus/Minus Query 9 CATCG-TTACGGCT-GGACTAC--GGGTATCTAATCCCTTTCGCTCCCCTAGCTTTCGTC 64 Sbjct 743 CATCGTTTACAGCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTC 684 Query 65 CCTGAGTGTCAGTTGCGGTCC-AGTAGAGAGCTTTCGCCACCGATGTTCTTCCCAATCTC 123 Sbjct 683 CCTGAGTGTCAGTTGCGG-CCTAGTAGAGCGCTTTCGCCACCGGTGTTCTTCCCAATCTC 625 Query 124 TACGCATTTCACCGCTACACTGGGAATTCCCTCTACCCCTACCGCACTCTAGCTATTCAG 183 Sbjct 624 TACGCATTTCACCGCTACACTGGGAATTCCCTCTACCCCTACCGCACTCTAGCTCTTCAG 565 Query 184 TTTCCACCGCCTACCCAGAGTTGAGCTCTGGTCTTTGACAGCAGACTTGAATAGCCACCT 243 Sbjct 564 TTTCCACCGCCTACCCGGAGTTAAGCTCCGGTCTTTGACAGCAGACTTGACGAGCCACCT 505 Query 244 GCGAACGCTTTACGCCCAATAATTCCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCT 303 Sbjct 504 GCGAACGCTTTACGCCCAATAATTCCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCT 445 Query 304 GCTGGCACGGAGTTAGCCGAGGCTTATTCCTCAGGTACCGTCAGATCTTCTTCCCTGAGA 363 Sbjct 444 GCTGGCACGGAGTTAGCCGAGGCTTATTCCTCAGGTACCGTCAGAACTTCTTCCCTGAGA 385 Query 364 AAAGAGGTTTACGATCCAAAGACCTTCTTCCCTCACGCGGTATTGCTCCGTCAGGCTTGC 423 Sbjct 384 AAAGAGGTTTACGATCCAAAGACCTTCTTCCCTCACGCGGTATTGCTCCGTCAGGCTTTC 325 Query 424 GCCCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTC 483
99 77 Sbjct 324 GCCCATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTC 265 Query 484 CCAGTGTGGCTGTTCTTCCTCTCAGAACAGCTACCGATCGCTGCCTTGGT-AGGCTTTTA 542 Sbjct 264 CCAGTGTGGCTGTTCTTCCTMTCAGAACAGCTACCGATCGCAGCCTTGGTGAGCCTTT-A 206 Query 543 CCCCACCAACTAGCTAATCGGACGCGAGCCCCTCTCTAGGCGATAAATC-TTTTACCCAA 601 Sbjct 205 CCCCACCAACTAGCTAATCGGACGCGAACCCCTCTCTAGGCGATAAATCCTTTTACCCGC 146 Query 602 AGGCGT-ATCGGGAATTAGCAGAAGTTTCCCTCTGTTGTCCCCGTCCTAGAGGCAGGTTC 660 Sbjct 145 AGGC-TCATCGGGAATTAGCAGAAGTTTCCCTCTGTTATCCCCGTCCTAGAGGCAGGTTT 87 Query 661 TCACGCGTTACTCACCCGTCCGCCACT-ATCTCCTAAGAGA-CCGTTCGACTTGCATGTG 718 Sbjct 86 TCACGTGTTACTCACCCGTCCGCCACTGAT-TCCTAAGA-ATCCGTTCGACTTGCATGTG 29 Query 719 TACAAAAATACCGCCAGCGTTCATCCTGA 747 Sbjct 28 TT-AAGCATACCGCCAGCGTTCATCCTGA H.Halospirulina sp. CCC Baja-95 Cl Halospirulina sp. MPI S3 MPI S3 Y Halospirulina tapeticola CCC Baja Usal 27F-809R Uncultured marine bacterium GU Spirulina subsalsa IAM M-223 AB Spirulina subsalsa AB2002/06 AY Spirulina subsalsa FACHB351 FJ Spirulina subsalsa AF Uip Geitlerinema sp. CCY S ribosomal RNA gene, partial sequence Length=1299 Score = 1033 bits (559), Expect = 0.0 Identities = 668/719 (92%), Gaps = 14/719 (1%) Strand=Plus/Plus Query 44 AGTGGCGGACGGGTGAGTAACGCGTGAGAATCTGCCTTCAGGACGGGGACAACAGCTGGA 103 Sbjct 1 AGTGGCGGACGGGTGAGTAACGCGTGAGAATCTACCTTCAGGACGGGGACAACAGCTGGA 60 Query 104 AACGGCTGCTAATACCCGATGTGCCGAGAGGTGAAATGTT-TTATCGCCTGAAGAGGAGC 162 Sbjct 61 AACGGCTGCTAATACCCGATATGCCGAAAGGTGAAAAGTTATT-TCGCCTGGAGAGGAGC 119 Query 163 TCGCGTCCGATTAGCTAGTTGGCGGGGTAAAAGCCCACCAAAGCGACGATCGGTAGCTGG 222 Sbjct 120 TCGCGTCCGATTAGCTAGTTGGTGGGGTAAAAGCTTACCAAGGCGACGATCGGTAGCTGG 179 Query 223 TCTGAGAGGACGAGCAGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCA 282 Sbjct 180 TCTGAGAGGATGAGCAGCCACACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCA 239 Query 283 GCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAACGCCGCGTGGGGGAAG 342 Sbjct 240 GCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAACGCCGCGTGGGGGAAG 299 Query 343 ACTGCCCTTGGGTTGTAAACCCCTTTTCTCAGGGAAG-AC-CCCCGACGGTACCTGAGGA 400
100 78 Sbjct 300 AAGGTTTTTGGATTGTAAACCCCTTTTCTCAAGGAAGAACAAAATGACGGTACTTGAGGA 359 Query 401 AGAAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTTATC 460 Sbjct 360 ATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTTATC 419 Query 461 CGGAATTATTGGGCGTAAAGCGTTCGTAGGCGGC-GTATCAAGTCTGCGGTCAAA-ATCC 518 Sbjct 420 CGGAATTATTGGGCGTAAAGCGTTCGTAGGCGGCTGT-TCAAGTCTGCTGTCAAAGA-CC 477 Query 519 GAGGCTCAACCTCGGGCCGGCCGTGGAAACTGAA-ACGCTAGAG-TACGGTAGGGGTAGA 576 Sbjct 478 GAGGCTCAACCTCGGGTCGGCAGTGGAAACTGAATA-GCTAGAGGT-CGGTAGGGGCAGA 535 Query 577 GGGAATTCCCGGTGTAGCGGTGAAATGCGTAGATATCGGGAAGAACACCGGTGGCGAAAG 636 Sbjct 536 GGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTGGGAAGAACACCGGTGGCGAAAG 595 Query 637 CGCTCTACTGGGCCGAAACTGACGCTGAGGAACGAAAGCTAGGGGAGCGAATGGGATTAG 696 Sbjct 596 CGCTCTGCTGGGCCGAACCTGACGCTGAGGGACGAAAGCTAGGGGAGCGAATGGGATTAG 655 Query 697 ATACCCCAGTAGTCCTAGCCGTAAACGATGGAGACTAGGTGTAGTCC-GTATCGACCCG 754 Sbjct 656 ATACCCCAGTAGTCCTAGCCGTAAACGATGGAAACTAGGTGTGG-CCTGTATCGACCCG 713 R Geitlerinema sp. CCY S ribosomal RNA gene, partial sequence Length=1299 Score = 990 bits (536), Expect = 0.0 Identities = 636/683 (93%), Gaps = 12/683 (1%) Strand=Plus/Minus Query 18 TACGGCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTTCCTCAGC 77 Sbjct 678 TACGGCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCCCTCAGC 619 Query 78 GTCAGTTTCGGCCCAGTAGAGCGCTTTCGCCACCGGTGTTCTTCCCGATATCTACGCATT 137 Sbjct 618 GTCAGGTTCGGCCCAGCAGAGCGCTTTCGCCACCGGTGTTCTTCCCAATATCTACGCATT 559 Query 138 TCACCGCTACACCGGGAATTCCCTCTACCCCTACCGTA-CTCTAGCGT-TTCAGTTTCCA 195 Sbjct 558 TCACCGCTACACTGGGAATTCCCTCTGCCCCTACCG-ACCTCTAGC-TATTCAGTTTCCA 501 Query 196 CGGCCGGCCCGAGGTTGAGCCTCGGAT-TTTGACCGCAGACTTGATAC-GCCGCCTACGA 253 Sbjct 500 CTGCCGACCCGAGGTTGAGCCTCGG-TCTTTGACAGCAGACTTGA-ACAGCCGCCTACGA 443 Query 254 ACGCTTTACGCCCAATAATTCCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCTGCTG 313 Sbjct 442 ACGCTTTACGCCCAATAATTCCGGATAACGCTTGCCTCCTCCGTATTACCGCGGCTGCTG 383 Query 314 GCACGGAGTTAGCCGAGGCTTCTTCCTCAGGTACCGTCGGGG-GT-CTTCCCTGAGAAAA 371 Sbjct 382 GCACGGAGTTAGCCGAGGCTGATTCCTCAAGTACCGTCATTTTGTTCTTCCTTGAGAAAA 323 Query 372 GGGGTTTACAACCCAAGGGCAGTCTTCCCCCACGCGGCGTTGCTCCGTCAGGCTTTCGCC 431 Sbjct 322 GGGGTTTACAATCCAAAAACCTTCTTCCCCCACGCGGCGTTGCTCCGTCAGGCTTTCGCC 263 Query 432 CATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCCA 491 Sbjct 262 CATTGCGGAAAATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGACCGTGTCTCAGTTCCA 203 Query 492 GTGTGGCTGCTCGTCCTCTCAGACCAGCTACCGATCGTCGCTTTGGTGGGCTTTTACCCC 551 Sbjct 202 GTGTGGCTGCTCATCCTCTCAGACCAGCTACCGATCGTCGCCTTGGTAAGCTTTTACCCC 143 Query 552 GCCAACTAGCTAATCGGACGCGAGCTCCTCTTCAGGCGATAA-AACATTTCACCTCTCGG 610 Sbjct 142 ACCAACTAGCTAATCGGACGCGAGCTCCTCTCCAGGCGA-AATAACTTTTCACCTTTCGG 84
101 79 Query 611 CACATCGGGTATTAGCAGCCGTTTCCAGCTGTTGTCCCCGTCCTGAAGGCAGATTCTCAC 670 Sbjct 83 CATATCGGGTATTAGCAGCCGTTTCCAGCTGTTGTCCCCGTCCTGAAGGTAGATTCTCAC 24 Query 671 GCGTTACTCACCCGTCCGCCACT 693 Sbjct 23 GCGTTACTCACCCGTCCGCCACT 1 Geitlerinema sp. CCY9619 GQ Phormidium sp. Ck3 GQ Oscillatoriales cyanobacterium Schier Phormidiaceae cyanobacterium SAG Phormidium sp. HBC9 EU Geitlerinema sp. CCY9412 GQ Phormidium sp. UTCC 487 AF Geitlerinema sp. CCY0102 GQ Uip 27F-809R 0.01 Çizelge 4.2: 27F-1492R Dizi analizi sonuçları Primerler Organizma Adı Maksimum Tanımlanma 1 CF (27F-1492R) U pse Oscillatoria rosea IAM M-220 gene for 16S ribosomal RNA % 96 1 CR (27F-1492R) U pse Oscillatoria rosea IAM M-220 gene for 16S ribosomal RNA % 96 2 CF (27F-1492R) U osc Uncultured cyanobacterium clone UFDCBA_30f 16S ribosomal RNA gene, % 96 partial sequence 2 CR (27F-1492R) U osc Uncultured cyanobacterium clone UFDCBA_30f 16S ribosomal RNA gene, % 93 partial sequence 3 CF (27F-1492R) KK Lyngbya birgei CCC S ribosomal RNA gene, partial sequence %98 3 CR (27F-1492R) KK Lyngbya birgei CCC S ribosomal RNA gene, partial sequence % 98 4 CF (27F-1492R) Bal pse Uncultured cyanobacterium clone Mat-CYANO-S19 16S ribosomal RNA %95 gene, partial sequence 4 CR (27F-1492R) Bal pse Uncultured cyanobacterium clone Mat-CYANO-S19 16S ribosomal RNA % 95 gene, partial sequence 5 CF(27F-1492R) ZD Geitlerinema sp. CR_13M 16S ribosomal RNA gene, partial sequence % 94 5 CR (27F-1492R) ZD Geitlerinema cf. acuminatum CCALA S ribosomal RNA gene, partial % 96 sequence 6 CF (27F-1492R) Şif Geitlerinema sp. PCC 7105 small subunit ribosomal RNA gene, partial % 96 sequence 6 CR (27F-1492R) Şif Geitlerinema sp. PCC 7105 small subunit ribosomal RNA gene, partial % 95 sequence 7 CF (27F-1492R) Bal ip Geitlerinema sp. PCC S rrna gene (partial), trna-ile gene, % 97 trna-ala gene and ITS1 and 23S rrna gene (partial), strain PCC CR (27F-1492R) Bal ip Geitlerinema sp. PCC S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 and 23S rrna gene (partial), strain PCC 8501 % 97 8 CF (27F-1492R) KY Phormidium animale NTMP03 16S ribosomal RNA gene, partial sequence % 97 8 CR (27F-1492R) KY Phormidium cf. terebriformis AB2002/07 16S ribosomal RNA gene, partial % 92 sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; 23S ribosomal RNA gene, partial sequence; and trna-ile and trna-ala genes, complete sequence 9 CF (27F-1492R) U sal Halospirulina sp.'ccc Baja-95 Cl.3', 16S rrna gene % 93 9 CR (27F-1492R)U sal Halospirulina sp.'ccc Baja-95 Cl.3', 16S rrna gene %91 10 CF (27F-1492R)U ip Geitlerinema sp. CCY S ribosomal RNA gene, partial sequence % CR (27F-1492R)U ip Geitlerinema sp. CCY S ribosomal RNA gene, partial sequence %91
102 80 Upse 27F-1492R Oscillatoria rosea IAM M-220 gene for 16S ribosomal RNA Length=1436 Score = 1589 bits (860), Expect = 0.0 Identities = 932/963 (96%), Gaps = 19/963 (1%) Strand=Plus/Plus Query 17 CATGC-AGTCG-ACG-AGTCTTCGGACTTAGTGGCGGACGGGTGAGTAACGCGTGAGAAT 73 Sbjct 26 CATGCAAGTCGAACGAAGTCTTCGGACTTAGTGGCGGACGGGTGAGTAACGCGTGAGAAT 85 Query 74 CTGCCTTCAGGATGGGGACAACAGTTGGAAACGACTGCTAATACCCAATATGCCTACGGG 133 Sbjct 86 CTGCCTTCAGGATGGGGACAACAGTTGGAAACGACTGCTAATACCCAATATGCCTACGGG 145 Query 134 TGAAACTTATATGGCCTGAAGATGAGCTCGCGTCTGATTAGCTAGTTGGTAGTGTAAGGG 193 Sbjct 146 TGAAACTTATTTGGCCTGAAGATGAGCTCGCGTCTGATTAGCTAGTTGGTGGTGTAAGAG 205 Query 194 ACTACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAG 253 Sbjct 206 ACTACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAG 265 Query 254 ACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCC 313 Sbjct 266 ACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAG-C 324 Query 314 TGACGGAGCAATACCGCGTGAGGGAGGAAGGTTTTTGGATTGTAAACCTCTTTTCTCAGG 373 Sbjct 325 TGACGGAGCAATACCGCGTGAGGGAGGAAGGTTTTTGGATTGTAAACCTCTTTTCTCAGG 384 Query 374 GAAGAATTTTGACGGTACCTGAGGAATAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGG 433 Sbjct 385 GAAGAATTTTGACGGTACCTGAGGAATAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGG 444 Query 434 TAATACGGAGGGTGCAAGCGTTATCCGGAATTATTGGGCGTAAAGAGTCCGTAGGTGGTC 493 Sbjct 445 TAATACGGAGGGTGCAAGCGTTATCCGGAATTATTGGGCGTAAAGAGTCCGTAGGTGGTC 504 Query 494 ATGCAAGTCTGCTGTCAAAGCCCACAGCTTAACTGTGGATCGGCGGTGGAAACTGCATGA 553 Sbjct 505 ATGCAAGTCTGCTGTCAAAGCCCACAGCTTAACTGTGGATAGGCGGTGGAAACTGCATGA 564 Query 554 CTTGAGTACGGTAGGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTGG 613 Sbjct 565 CTTGAGTACGGTAGGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTGG 624 Query 614 GAAGAACACCGGTGGCGAAAGCGCTCTACTGGACCGTAACTGACACTGAGGGACGAAAGC 673 Sbjct 625 -AAGAACACCGGTGGCGAAAGCGCTCTACTGGACCGTAACTGACACTGAGGGACGAAAGC 683 Query 674 CAAGGTAGCGAAAGGGATTAGATACCCCTGTAGTCTTGGCCGTAAACGATGGATACTAGG 733 Sbjct 684 CAAGGTAGCGAAAGGGATTAGATACCCCTGTAGTCTTGGCCGTAAACGATGGATACTAGG 743 Query 734 CGTTGTCTGTATCGACCCGGACAGTGCCGAAGCTAACGCGTTAAGTATCCCGCCTGGGGA 793 Sbjct 744 CGTTGTCTGTATCGACCCGGACAGTGCCGAAGCTAACGCGTTAAGTATCCCGCCTGGGGA 803 Query 794 GTACGCACGCAAGTGTGAAACTCAA-GGAATTGACGGGGGCCCGCACAAGCGGTGGAGTA 852 Sbjct 804 GTACGCACGCAAGTGTGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGTA 863 Query 853 TGTGGTT-AATTTCGATGCA-CGCGGAGAGC-TTACCTAGATTTGACATCC-GTGA-CT- 906 Sbjct 864 TGTGGTTTAATT-CGATGCAACGCGAAGAACCTTACCAAGATTTGACATCCCGTGAACTA 922 Query 907 TC-AGAGATG-ATTGCTGCCTTCGGTA-CACGCAGACATGTTGCGTGCATG-CTGTCGTC 962 Sbjct 923 TCCAGAGATGGATGGGTGCCTTCGGGAACACGGAGACAGGT-G-GTGCATGGCTGTCGTC 980 Query 963 AGC 965
103 81 Sbjct 981 AGC 983 R Oscillatoria rosea IAM M-220 gene for 16S ribosomal RNA Length=1436 Score = 1604 bits (868), Expect = 0.0 Identities = 940/971 (96%), Gaps = 19/971 (1%) Strand=Plus/Minus Query 13 GCCTTCGGCG-TCTCC-CCGCG-ACGGTTAGAGATAACGACTTCGGGCGTTGCCAGCTTC 69 Sbjct 1388 GCCTTCGGCGCCCTCCTCCGCGAACGGTTAGAG-TAACGACTTCGGGCGTTGCCAGCTTC 1330 Query 70 CATGGTGGGACGGGGGGTGTGTACAAGGCCCGGGAACGGATTCACCGCATTATGCTGACC 129 Sbjct 1329 CATGGTGTGACGGGCGGTGTGTACAAGGCCCGGGAACGGATTCACCGCAGTATGCTGACC 1270 Query 130 TGCGATTACTAGCGATTCCTCCTTCATGCAGGCGAGTTGCAGCCTGCAATCTGAACTGAG 189 Sbjct 1269 TGCGATTACTAGCGATTCCTCCTTCATGCAGGCGAGTTGCAGCCTGCAATCTGAACTGAG 1210 Query 190 GCCGGGTTTGATGAGATTCGCTTGCCCTCGCGGGGTTGCTGCCCTTTGTCCCGACCATTG 249 Sbjct 1209 GCCGGGTTTGATGAGATTCGCTTGCCCTCGCGGGGTTGCTGCCCTTTGTCCCGACCATTG 1150 Query 250 TAGTACGTGTGTAGCCCAAAATGTAAGGGGCATGATGACTTGACGTCATCCCCACCTTCC 309 Sbjct 1149 TAGTACGTGTGTAGCCCAAAATGTAAGGGGCATGATGACTTGACGTCATCCCCACCTTCC 1090 Query 310 TCCGAGTTGTCCCCGGCAGTCTCCCTAGAGTGCCCAACTTAATGATGGCAACTAAGGACG 369 Sbjct 1089 TCCGAGTTGTCCCCGGCAGTCTCCCTAGAGTGCCCAACTTAATGATGGCAACTAAGGACG 1030 Query 370 AGGGTTGCGCTCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCA 429 Sbjct 1029 AGGGTTGCGCTCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCA 970 Query 430 TGCACCACCTGTCTCCGTGTTCCCGAAGGCACCAATCCATCTCTGGAAAGTTCACGGGAT 489 Sbjct 969 TGCACCACCTGTCTCCGTGTTCCCGAAGGCACCCATCCATCTCTGGATAGTTCACGGGAT 910 Query 490 GTCAAATCTTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTG 549 Sbjct 909 GTCAAATCTTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTG 850 Query 550 TGCGGGCCCCCGTCAATTCCTTTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCGGGAT 609 Sbjct 849 TGCGGGCCCCCGTCAATTCCTTTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCGGGAT 790 Query 610 ACTTAACGCGTTAGCTTCGGCACTGTCCGGGTCGATACAGACAACGCCTAGTATCCATCG 669 Sbjct 789 ACTTAACGCGTTAGCTTCGGCACTGTCCGGGTCGATACAGACAACGCCTAGTATCCATCG 730 Query 670 TTTACGGCCAAGACTACAGGGGTATCTAATCCCTTTCGCTACCTTGGCTTTCGTCCCTCA 729 Sbjct 729 TTTACGGCCAAGACTACAGGGGTATCTAATCCCTTTCGCTACCTTGGCTTTCGTCCCTCA 670 Query 730 GTGTCAGTTACGGTCCAGTAGAGCGCTTTCGCCACCGGTGTTCTTCCCAATATCTACGCA 789 Sbjct 669 GTGTCAGTTACGGTCCAGTAGAGCGCTTTCGCCACCGGTGTTCTTCC-AATATCTACGCA 611 Query 790 TTTCACCGCTACACTGGGAATTTCCCTCTACCCCTACCGTACTCAAGTCATGCAGTTTCC 849 Sbjct 610 TTTCACCGCTACACTGGGAATT-CCCTCTACCCCTACCGTACTCAAGTCATGCAGTTTCC 552 Query 850 ACCGCCGATCCACAGATAAGCTGTGGCCTTTGACAGCCAGACTAGCATGGACCACCCTTA 909 Sbjct 551 ACCGCCTATCCACAGTTAAGCTGTGGGCTTTGACAGC-AGACTTGCATG-ACCACC-T-A 496 Query 910 CGGACTCTTTACGCC-AATA-TGCAGGATA-CGCTTGCCACC-TC-GTAT-ACCGCGGCT 963
104 82 Sbjct 495 CGGACTCTTTACGCCCAATAATTCCGGATAACGCTTGC-ACCCTCCGTATTACCGCGGCT 437 Query 964 GCCTGGCCACG 974 Sbjct 436 GC-TGGC-ACG Synechococcus sp. PCC 7117 AB Phormidium angustissimum KMD34 EF Synechococcus elongatus CCMP1630 AY94... Synechococcus sp. PCC 8807 AF U pse 27F-1494R 77 Oscillatoria rosea IAM M-220 AB Uncultured Oscillatoria sp. EF Synechococcus sp. Salton Sea DQ Phormidium sp. OL S4 AM Phormidium sp. OL 81 AM Phormidium sp. MBIC10210 AB Synechococcus sp. UH7 AF Leptolyngbya sp. CR L33 EF U osc Uncultured cyanobacterium clone UFDCBA_30f 16S ribosomal RNA gene, partial sequence Length=1454 Score = 1450 bits (785), Expect = 0.0 Identities = 857/889 (96%), Gaps = 15/889 (1%) Strand=Plus/Plus Query 15 ACATGC-AGTCG-ACG-ACTCTTCGGTAGCTGAGTGCGCGGACGGGTGAGTAACGCGTGA 71 Sbjct 47 ACATGCAAGTCGAACGAACTCTTCGG-AG-TGAGTG-GCGGACGGGTGAGTAACGCGTGA 103 Query 72 GAATCTGCCTGTGGGTCGGGGATAACCACTGGAAACGGTGGCTAATACCGGATGAGCCGA 131 Sbjct 104 GAATCTGCCTGTGGGTCGGGGACAACTACTGGAAACGGTGGCTAATACCGGATGAGCCGA 163 Query 132 AAGGTAAAAGATTTATTGCCCACAGATGAGCTCGCGTCTGATTAGCTAGTTGGTGTGGTA 191 Sbjct 164 AAGGTAAAAGATTTATTGCCCACAGATGAGCTCGCGTCTGATTAGCTAGTTGGTAAGGTA 223 Query 192 AAAGCGCACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGAC 251 Sbjct 224 AAAGCGTACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGAC 283 Query 252 TGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAA 311 Sbjct 284 TGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAA 343 Query 312 AGCCTGACGGAGCAAGACCGCGTGGGGGAGGAAGGTTTTTGGATCGTAAACCCCTTTTGT 371 Sbjct 344 AGCCTGACGGAGCAAGACCGCGTGGGGGAGGAAGGTTTTTGGATCGTAAACCCCTTTTGT 403 Query 372 CAAGGAAGAAGCTCTGACGGTACTTGACGAATAAGCCTCGGCTAACTCCGTGCCAGCAGC 431 Sbjct 404 CAAGGAAGAAGCTCTGACGGTACTTGACGAATAAGCCTCGGCTAACTCCGTGCCAGCAGC 463 Query 432 CGCGGTAATACGGAGGAGGCAAGCGTTATCCGGAATAATTGGGCGTAAAGAGTCTGTAGG 491 Sbjct 464 CGCGGTAATACGGAGGAGGCAAGCGTTATCCGGAATGATTGGGCGTAAAGCGTCTGTAGG 523
105 83 Query 492 TGGCGAACTAAGTCTGCTGTTAAAGATCACAGCTCAACTGTGGGTAGGCAGTGGAAACTG 551 Sbjct 524 TGGCCAGCTAAGTCTGCTGTTAAAGATCACAGCTCAACTGTGGGAAGGCAGTGGAAACTG 583 Query 552 GTTAGCTAGAGGCTGGTAGGGGTAAAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGAT 611 Sbjct 584 GCAAGCTAGAGGTTGGTAGGGGTAAAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGAT 643 Query 612 ATCGGGAAGAACACCGGTGGCGAAAGCGCTTTACTGGGCCAGACCTGACACTGAAGGACG 671 Sbjct 644 ATCGGGAAGAACACCGGTGGCGAAAGCGCTTTACTGGGCCAGACCTGACACTGAAGGACG 703 Query 672 AAAGCTAGGGGAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCCGTAA-CGATGGAGA 730 Sbjct 704 AAAGCTAGGGGAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCTGTAAACGATGGAGA 763 Query 731 CTAGGTGTAGCACGTATCGACCCGTGCTGTATCGTAGCCAACGCGTTAAGTCTCCCGCCT 790 Sbjct 764 CTAGGTGTAGCACGTATCGACCCGTGCTGTATCGTAGCCAACGCGTTAAGTCTCCCGCCT 823 Query 791 GGGGAGTACGCACGCAAGTGTGAAACTCAAAGGAATTTGACGGGGGCCCGCACAAGCGGT 850 Sbjct 824 GGGGAGTACGCACGCAAGTGTGAAACTCAAAGGAATT-GACGGGGGCCCGCACAAGCGGT 882 Query 851 GGAGTATGTGGGATTTTAATTTCGATGCCATCGCAGAGGACCCTTTACC 899 Sbjct 883 GGAGTATGTGG--TTT-AATT-CGATGCAA-CGC-GAAGAACCTT-ACC 924 R Uncultured cyanobacterium clone UFDCBA_30f 16S ribosomal RNA gene, partial sequence Length=1454 Score = 1181 bits (639), Expect = 0.0 Identities = 764/820 (93%), Gaps = 25/820 (3%) Strand=Plus/Minus Query 38 TTAGGGTAACGACTTCGGGCGTGGCCAACTTCCATGGGGTGACGGGAGGGGGGAACAAGG 97 Sbjct 1385 TTAGGGTAACGACTTCGGGCGTGGCCAACTTCCATGGTGTGACGGGCGGTGTGTACAAGG 1326 Query 98 CCGGGGAACGGATTCACCGCCGGATGCTGACCGGCGATTACTAGCGATTCCTCCTTCATG 157 Sbjct 1325 CCCGGGAACGGATTCACCGCCGTATGCTGACCGGCGATTACTAGCGATTCCTCCTTCATG 1266 Query 158 CTCTCGAGTTGAAGAGAGCAATCTGAACTGAGACCGGGTTTCTGGGATTAGCTGGCCC-T 216 Sbjct 1265 CTCTCGAGTTGCAGAGAGCAATCTGAACTGGGACCGGGTTTCTGAGATTAGCT-TCCCTT 1207 Query 217 CGCGGGTTCGCTGCCCTTTGTCCCGACCATTGTAGTACGTGTGTCGCCCAGGGCGTAAGG 276 Sbjct 1206 CGCAGGTTCGCTGCCCTTTGTCCCGACCATTGTAGTACGTGTGTTGCCCAGGGCGTAAGG 1147 Query 277 GGCATGCTGACTTGACGTCATCCCCACCTTCCTCCGGTTTGTCACCGGCAGTCTCTCTAG 336 Sbjct 1146 GGCATGCTGACTTGACGTCATCCCCACCTTCTTCCGGTTTGTCACCGGCAGTCTCTCTAG 1087 Query 337 AGTTCCCT-ACTTAA-AGCTGGCAACTAAAGACGAGGGTTGCGCTCGTTGCGGGACTTAA 394 Sbjct 1086 AGTT-CCTAACTTAATA-TTGGCAACTAAAGACGAGGGTTGCGCTCGTTGCGGGACTTAA 1029 Query 395 CCCAACATCTCACGACACGAGCTGACGACAGCCATGCACCACCTGTGTTCGCGCTCCCGA 454 Sbjct 1028 CCCAACATCTCACGACACGAGCTGACGACAGCCATGCACCACCTGTGTTCGCGCTCCCGA 969 Query 455 AGGCACTTTCAAGTTTCCCTGAA-ATTCGCGACATGTCAAGCCCTGGTAAGGTTCTTCGC 513 Sbjct 968 AGGCACTCTCAACTTTCA-TCAAGATTCGCGACATGTCAAGCCCTGGTAAGGTTCTTCGC 910 Query 514 GTTGCATCAAATTAAACCACATACTCCACCGCTTGTGCGGGCCCCCGTCAATTCTTTTGA 573 Sbjct 909 GTTGCATCGAATTAAACCACATACTCCACCGCTTGTGCGGGCCCCCGTCAATTCCTTTGA 850
106 84 Query 574 GTTTCACACTTGCGGTGCGTACTCCCCAGGCGGGAGACTTAACGCGTTGGCTACGATACA 633 Sbjct 849 GTTTCACACTTGCG-TGCGTACTCCCCAGGCGGGAGACTTAACGCGTTGGCTACGATACA 791 Query 634 GACACGGGTCGATACGTGCTACACCTAGTCTCCATCGTTTACGGGCTAGAACTACAGGGG 693 Sbjct 790 G-CACGGGTCGATACGTGCTACACCTAGTCTCCATCGTTTACAG-CTAGGACTACAGGGG 733 Query 694 GTATCTAATCCCTTTCGCCTCCCCTAGCTTTCGTCCTTCAGTGTCAGGTCTAGACATAGT 753 Sbjct 732 -TATCTAATCCCTTTCGC-TCCCCTAGCTTTCGTCCTTCAGTGTCAGGTCTGGCCC-AGT 676 Query 754 AAAGCTGCTTTTCGCGACCGGTTGATTCTTCTAGATATCTACGCAATTTCACCGCTACAG 813 Sbjct 675 AAAGC-GCTTT-CGCCACCGGT-G-TTCTTCCCGATATCTACGCA-TTTCACCGCTACAC 621 Query 814 CGGGGTAATTCACCTTTTACCCCATACCAGCCATCTAGCT 853 Sbjct 620 CGGG--AATTC-CCTTT-ACCCC-TACCAACC-TCTAGCT Pseudoscillatoria coralii BgP10 4S FJ... Uncultured cyanobacterium clone BB2IN... Uncultured cyanobacterium clone UFDCB... Uncultured cyanobacterium clone UNFR... U osc 27F-1492R KK Lyngbya birgei CCC S ribosomal RNA gene, partial sequence Length=706 Score = 1253 bits (678), Expect = 0.0 Identities = 697/706 (98%), Gaps = 1/706 (0%) Strand=Plus/Plus Query 28 GTGTCTTCGGACACAGTGGCGGACGGGTGAGTAACGCGTGAGAATCTACCTTCAGGTCGG 87 Sbjct 1 GTGTCTTCGGACACAGTGGCGGACGGGTGAGTAACGCGTGAGAATCTACCTTCAGGTCGG 60 Query 88 GGATAACCCCTGGAAACGGGGGCTAATACCGGATAGGTCGAAAGATGAAAGGTTAACTGC 147 Sbjct 61 GGATAACCCCTGGAAACGGGGGCTAATACCGGATAGGTCGAAAGATGAAAGGTTAACTGC 120 Query 148 CTGAAGAGGAGCTCGCGTCTGATTAGCTAGTAGGTGTGGTAAGAGCGCACCTAGGCGACG 207 Sbjct 121 CTGAAAAGGGGCTCGCGTCTGATTAGCTAGTAGGTGTGGTAAGAGCGCACCTAGGCGACA 180 Query 208 ATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCCAGACTC 267 Sbjct 181 ATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCCAAACTC 240 Query 268 CTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACC 327 Sbjct 241 CTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACC 300 Query 328 GCGTGGGGGAGGAAGGCCTTTGGGTTGTAAACCCCTTTTGTTAGGGAAGaaaaaaaTGAC 387 Sbjct 301 GCGGGGGGGAGGAAGGCCTTTGGGTTGTAAACCCCTTTTGTTAGGGAAAAAAAAAATGAC 360 Query 388 GGTACCTAACGAATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAG 447 Sbjct 361 GGTACCTAACGAATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAG 420 Query 448 GCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGCCCGGCAAGTCTGCT 507 Sbjct 421 GCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGCCCGGCAAGTCTGCT 480
107 85 Query 508 GTCAAAGACCACAGCTCAACTGTGGAAAGGCAGTGGAAACTGCAAGGCTAGAGTGCGGTA 567 Sbjct 481 GTCAAAGACCACAGCTCAACTGTGGAAAGGCAGTGGAAACTGCAAGGCTAGAGTGCGGTA 540 Query 568 GGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGAGATTGGGAAGAACACCGGT 627 Sbjct 541 GGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGAGATTGGAAAGAACACCGGT 600 Query 628 GGCGAAAGCGCTCTACTGGGCCGCAACTGACACTGATGGACGAAAGCTAGGGGAGCGAAT 687 Sbjct 601 GGCGAAAGCGCTCTACTGGGCCGCAACTGACACTGATGGACGAAAGCTAGGGGAGCGAAT 660 Query 688 GGGATTAGATACCCC-AGTAGTCCTAGCTGTAAACGATGGAAACTA 732 Sbjct 661 GGGATTAGATCCCCCCAGTAGTCCTAGCTGTAAACGATGGAAACTA 706 Oscillatoria spongeliae 33P1 AY Oscillatoria spongeliae 35P1 AY Oscillatoria spongeliae 39P1 AY Oscillatoria spongeliae 310P1 AY Microcoleus chthonoplastes CCY0602 GQ... KK 27F-1492R 100 Lyngbya birgei CCC 333 EU Bal pse Uncultured cyanobacterium clone Mat-CYANO-S19 16S ribosomal RNA gene, partial sequence Length=1413 Score = 893 bits (483), Expect = 0.0 Identities = 538/563 (95%), Gaps = 10/563 (1%) Strand=Plus/Plus Query 18 TGC-AGTCG-ACGGAGTGCTTTCGGAGCACTTAGTGGCGGACGGGTGAGTAACGCGTGAG 75 Sbjct 29 TGCAAGTCGAACGGAGTGCCTTCGG-GCACTTAGTGGCGGACGGGTGAGTAACGCGTGAG 87 Query 76 AATCTG-CCTTCAGGTCGGGGATAACGGCTGGAAACGGTCGCTAATACCGGATGTGCCGA 134 Sbjct 88 GATCTGCCCTT-AGGTCGGGGATAACGACTGGAAACGGTCGCTAATACCGGATATGCCGA 146 Query 135 GAGGTGAAAGGTTAACTGCCTGAGGATGAGCTCGCGTCGGATTAGCTAGTTGGTGGGGTA 194 Sbjct 147 GAGGTGAAAGGTTAACTGCCTGAGGATGAGCTCGCGTCGGATTAGCTAGTTGGTGGGGTA 206 Query 195 AGGGCCTACCAAGGCGACGATCCGTAGCTGGTTTGAGAGGATGACCAGCCACACTGGGAC 254 Sbjct 207 AGGGCCTACCAAGGCGACGATCCGTAGCTGGTTTGAGAGGATGACCAGCCACACTGGGAC 266 Query 255 TGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGCA 314 Sbjct 267 TGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGCA 326 Query 315 AGCCTGACGGAGCAACGCCGCGTGCGGGAGGAAGGCCTGTGGGTTGTAAACCGCTTTTCT 374 Sbjct 327 AGCCTGACGGAGCAACGCCGCGTGGGGGAGGAAGGCCTGTGGGTTGTAAACCTCTTTTCT 386 Query 375 CAAGGAAGAAGTCAGTGACGGTACTTGAGGAATCAGCATCGGCTAACTCCGTGCCAGCAG 434 Sbjct 387 CAAGGAAGAAGCCAGTGACGGTACTTGAGGAATCAGCATCGGCTAACTCCGTGCCAGCAG 446 Query 435 CCGCGGTAAGACGGAGGATGCAAGCGTTATCCGGAATGATTGGGCGTAAAGCGTCCGCAG 494
108 86 Sbjct 447 CCGCGGTAAGACGGAGGATGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGCAG 506 Query 495 GTGGTTATACAAGTCTGCCGTTAAAGCGTGGGGCTTAACTCCTTAAgggggggggAAAAC 554 Sbjct 507 GTGGTTATACCAGTCTGCCGTTAAAGCGTGGG-CTTAACTCCATAAAGGCGGTG-AAA-C 563 Query 555 TGTCTGACAA-AGTGAGG-AGGG 575 Sbjct 564 TGTATGACTAGAGTGAGGTAGGG 586 R Uncultured cyanobacterium clone Mat-CYANO-S19 16S ribosomal RNA gene, partial sequence Length=1413 Score = 1507 bits (816), Expect = 0.0 Identities = 914/958 (95%), Gaps = 20/958 (2%) Strand=Plus/Minus Query 14 CTTCGGC-GT-CCCCTCC-GGA-AGGTTGGAGAAACGACTTCGGGCATGGCCAACTCCCA 69 Sbjct 1392 CTTCGGCGGTCCCCCTCCATTACTGGTTGGAGTAACGACTTCGGGCATGGCCAACTCCCA 1333 Query 70 TGGTGTGACggggggggTGTACAAGGCCCGGGAACGTATTCACCGCCGTATGCTGACCGG 129 Sbjct 1332 TGGTGTGACGGGCGGTGTGTACAAGGCCCGGGAACGTATTCACCGCCGTATGCTGACCGG 1273 Query 130 CGATTACTAGCGATTCCGCCTTCATGCAGGCGAGTTGCAGCCTGCAATCTGAACTGGGCC 189 Sbjct 1272 CGATTACTAGCGATTCCTCCTTCATGCAGGCGAGTTGCAGCCTGCAATCTGAACTGGGCC 1213 Query 190 ACGGTTTACGAGATTAGCTCACCCTCGCGGACTGGCAACCCTCTGTCCGTGGCATTGTAG 249 Sbjct 1212 ACGGTTTACGAGATTGGCTTACCCTCGCGGACTCGCTACCCTCTGTCCGTGGCATTGTAG 1153 Query 250 TACGTGTGTAGCCCAAAATGTAAGGGGCATGCTGACTTGACGTCATCCACACCTTCCTCC 309 Sbjct 1152 TACGTGTGTAGCCCAGAATGTAAGGGGCATGCTGACTTGACGTCATCCACACCTTCCTCC 1093 Query 310 GGTTTGTCACCGGCAGTCTCTCTAGAGTGCCCAACTGAATGCTGGCAACTAAAAACGTGG 369 Sbjct 1092 GGTTTGTCACCGGCAGTCTCTCTAGAGTGCCCAACTGAATGCTGGCAACTAAAAACGTGG 1033 Query 370 GTTGCGCTCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCATGC 429 Sbjct 1032 GTTGCGCTCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCATGC 973 Query 430 ACCACCTGTCTCCCCGCTCCCGTAGGCACTCCCCCCTTTCAGGAGGATTCGGGGGATGTC 489 Sbjct 972 ACCACCTGTCTCCCCGCTCCCGTAGGCACTCCTCCCTTTCAAGAGGATTCGGGGGATGTC 913 Query 490 AAACCCTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTGC 549 Sbjct 912 AAACCCTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTGC 853 Query 550 GGGCCCCCGTCAATTCCTTTGAGTTTCACTCTTGCGAGCGTACTCCCCAGGCGGGATACT 609 Sbjct 852 GGGCCCCCGTCAATTCCTTTGAGTTTCACTCTTGCGAGCGTACTCCCCAGGCGGGATACT 793 Query 610 TAACGCGTTAGCTTCGGCACGGCCCGGGTCGATACGAACCACACCTAGTATCCATCGTTT 669 Sbjct 792 TAACGCGTTAGCTTCGGCACTGTCCGGGTCGATACGAACAACACCTAGTATCCATCGTTT 733 Query 670 ACGGCTAGGACTACAGGGGTATCTAATCCCTTTCGCTCCCCTAGCTTTCGTCCCTCAGTG 729 Sbjct 732 ACGGCTAGGACTACAGGGGTATCTAATCCCTTTCGCTCCCCTAGCTTTCGTCCCTCAGTG 673 Query 730 TCAGTCAAGGCCCAGTAGAGCGCTTTCGCCACTGGTGTTCTTCCCGATATCTACGCATTT 789 Sbjct 672 TCAGTCAAGGCCCAGTAGAGCGCTTTCGCCACTGGTGTTCTTCCCGATATCTACGCATTT 613 Query 790 CACCGCTACACCGGGAATTCCCTCTACCCCTACCTCACTCTAGTCATACAGTTCCACCCG 849 Sbjct 612 CACTGCTACACCGGGA-TTCCCTCTACCCCTACCTCACTCTAGTCATACAGTTTCACC-G 555
109 87 Query 850 CCCTTATGGAGTTAAGCCCCCACGCTTTA-CGGCAGACTTGTTATAACCACCTGCG-ACG 907 Sbjct 554 CCTTTATGGAGTTAAGCCC--ACGCTTTAACGGCAGACTGGT-ATAACCACCTGCGGACG 498 Query 908 CTT-ACGCCCAATCCAT-CAG-ATA-CGCTGGCATCCCTCCGATCTTAACCGCCGGCT 961 Sbjct 497 CTTTACGCCCAATA-ATTCCGGATAACGCTTGCATCC-TCCG-TCTTA-CCGC-GGCT Leptolyngbya badia CRS-1 EF Leptolyngbya badia CRS-1 EF Leptolyngbya sp. JO2-1B EU Leptolyngbya badia CRS-1 EF Uncultured cyanobacterium clone Mat-C... Leptolyngbya antarctica ANT.LH67.1 AY... Leptolyngbya sp. FYG FJ Bal pse 27F-1492R 0.1 ZD Geitlerinema sp. CR_13M 16S ribosomal RNA gene, partial sequence Length=1181 Score = 1373 bits (743), Expect = 0.0 Identities = 843/890 (94%), Gaps = 15/890 (1%) Strand=Plus/Plus Query 82 CAGGAGGGGGACAACCACTGGAAACGGTGGCTAATACCCCATAAGCCGAGAGGTAAAAAG 141 Sbjct 1 CAGGAGGGGGACAACCATTGGAAACGGTGGCTAATACCCCATAAGCCGAGAGGTGAAAAG 60 Query 142 GAATCCGCCTGTGGAGGAGCTCGCGTCTGATTAGCTAGTAGGTGAGGTAAGAGCTTACCT 201 Sbjct 61 GAATCCGCCTGTGGAGGAGCTCGCGTCTGATTAGCTAGTAGGTAAGGTAAAAGCTTACCT 120 Query 202 AGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGC 261 Sbjct 121 AGGCGACGATCAGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGC 180 Query 262 CCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGA 321 Sbjct 181 CCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGA 240 Query 322 GCCAGACCGCGTGAGGGAAGAAGGCCTTTGGGTTGTAAACCTCTTTTCTCAGGGAAGAAG 381 Sbjct 241 GCCAGACCGCGTGAGGGAAGAAGGCCCTTGGGTTGTAAACCTCTTTTCTCAAGGAAGAAG 300 Query 382 AAAGTGACGGTACCTGAGG-AATCAG-CCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAA 439 Sbjct 301 CAAGTGACGGTACTTGAGGCAAT-AGGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAA 359 Query 440 GACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGTGATT 499 Sbjct 360 GACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGTGACT 419 Query 500 CAAGTCTGCTGTCAAAACCCAGGGCTTAACTCTGGATCGGCAGTGGAAACTGAATGACTA 559 Sbjct 420 CAAGTCTGCTGTCAAAAACTAGGGCTTAACTCTAGAGAGGCAGTGGAAACTGAGTAACTA 479 Query 560 GAGTGCGGCAGGGGCAGAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGAGATCGGGAA 619 Sbjct 480 GAGAGCGGCAGGGGCAGAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGRGATCGGGAA 539 Query 620 GAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGCGACTGACACTCACAGGACGAAAGCTA 679
110 88 Sbjct 540 GAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGCGACTGACACTCACAGGACGAAAGCTA 599 Query 680 GGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCTGTAAACGATGGAAACTAGGTG 739 Sbjct 600 GGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCCGTAAACGATGGAAACTAGGTG 659 Query 740 TGGCCTGTATCGACCCGGGCCGTGCCGGAGCTAACGCGTTAAGTTTCCCGCCTGGGGAGT 799 Sbjct 660 TGGCCTGTATCGACCCGGGCCGTGCCGGAGCTAACGCGTTAAGTTTCCCGCCTGGGGAGT 719 Query 800 ACGCACGCAAGTGTGAAACTCAAGGGAATTTGACGGGGGCCCGCACAAGCGGTGGGAGTA 859 Sbjct 720 ACGCACGCAAGTGTGAAACTCAAAGGAATT-GACGGGGGCCCGCACAAGCGGTGG-AGTA 777 Query 860 TGTGGGTTTAATTCGATGCAACGGCGGAGAACCTTAC-AGT--CTTGACATGGTCGCGAA 916 Sbjct 778 TGTGG-TTTAATTYGATGCAACG-CGAAGAACCTTACCAGGGACTTGACATG-TCGCGAA 834 Query 917 TCCCGGTTGAAGGCTGGGCAGTGACGTCGGAAGCGCGTAACACAGGATGG 966 Sbjct 835 TCTTGGG-GAAACCTGAG-AGTGCCTTCGGGAGCGCG-AACACAGG-TGG 880 R Geitlerinema cf. acuminatum CCALA S ribosomal RNA gene, partial sequence Length=1165 Score = 1535 bits (831), Expect = 0.0 Identities = 914/951 (96%), Gaps = 18/951 (1%) Strand=Plus/Minus Query 18 TTCGGGCGTCCTCCTCCGCG-ACGGTTAGAAGTAAACGACTTCGGGCGTGGCCAGCTCCC 76 Sbjct 1092 TTC-GGCGTCCTCCTCCGCGAACGGTTAG-AGT-AACGACTTCGGGCGTGGCCAGCTCCC 1036 Query 77 ATGGTGTGACGGGGGGTGTGTACAAGGCCCGGGAACGGATTCACTGCCGTATGCTGACCG 136 Sbjct 1035 ATGGTGTGACGGGCGGTGTGTACAAGGCCCGGGAACGGATTCACTGCCGTATGCTGACCG 976 Query 137 GCAATTACTAGCGATTCCTCCTTCACGCAGGCGAGTTGCAGCCTGCGATCTGAACTGTGG 196 Sbjct 975 GCAATTACTAGCGATTCCTCCTTCACGCAGGCGAGTTGCAGCCTGCGATCTGAACTGTGG 916 Query 197 CTGGGTTTATGGGATTGGCTCGTCCTCGCGGCTTGGCTACCCTTTGTCCCAACCATTGTA 256 Sbjct 915 CTGAGTTTATGGGATTGGCTTGTCCTTGCGGCTTAGCGACCCTTTGTCCCAACCATTGTA 856 Query 257 GTACGTGTGTCGCCCAGGACGTAAGGGGCATGCTGACTTGACGTCATCCCCACCTTCCTC 316 Sbjct 855 GTACGTGTGTAGCCCAGGACGTAAGGGGCATGCTGACTTGACGTCATCCCCACCTTCCTC 796 Query 317 CGGTTTGTCACCGGCAGTCTCCCTAGAGTTCCCATCTGAATGCTGGCAACTAAGGACGAG 376 Sbjct 795 CGGTTTGTCACCGGCAGTCTCCCTAGAGTTCCCATCTGAATGCTGGCAACTAAGGACGAG 736 Query 377 GGTTGCGCTCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCATG 436 Sbjct 735 GGTTGCGCTCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCATG 676 Query 437 CACCACCTGTGTTCGCGCTCCCGAAGGCACTCCCAGCTTTCACCGGGATTCGCGACATGT 496 Sbjct 675 CACCACCTGTGTTCGCGCTCCCGAAGGCACTCTTAGGTTTCCCCAAGATTCGCGACATGT 616 Query 497 CAAGCCCTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTG 556 Sbjct 615 CAAGCCCTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTG 556 Query 557 CGGGCCCCCGTCAATTCCTTTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCGGGAAAC 616 Sbjct 555 CGGGCCCCCGTCAATTCCTTTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCGGGAAAC 496 Query 617 TTAACGCGTTAGCTCCGGCACGGCCCGGGTCGATACAGGCCACACCTAGTTTCCATCGTT 676
111 89 Sbjct 495 TTAACGCGTTAGCTCCGGCACGGCCCGGGTCGATACAGGCCACACCTAGTTTCCATCGTT 436 Query 677 TACAGCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCCTGTGAG 736 Sbjct 435 TACGGCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCCTGTGAG 376 Query 737 TGTCAGTCGCGGCCCAGCAGAGCGCTTTCGCCACCGGGTGTTCTTCCCGATCTCTACGCA 796 Sbjct 375 TGTCAGTCGCGGCCCAGCAGAGCGCTTTCGCCACCGG-TGTTCTTCCCGATCTCTACGCA 317 Query 797 TTTTCACCGCTACACCGGGAAATTCCCTCTGCCCCTGCCGCACTCTAGTCATTCAGTTTT 856 Sbjct 316 TTT-CACCGCTACACCGGGAA-TTCCCTCTGCCCCTGCCGCTCTCTAGTCACTCAGTTT- 260 Query 857 CCCACTGCCGATCCAGAGTTAAGCCCTTGGGTTTTGACAGCAGACTTGAATCACCACCTA 916 Sbjct 259 CC-ACTGCCGATCCAGAGTTAAGCCCT-GGGTTTTGACAGCAGACTTGAATAACCACCTA 202 Query 917 CGGACGCTT-ACGCCGAAATA-TTCCAGAATA-CGTCTTGGCCTCCCTCCG 964 Sbjct 201 CGGACGCTTTACGCCCAA-TAATTCCGGA-TAACG-CTTG-CCTCC-TCCG Microcoleus chthonoplastes SAG 2209 E... Microcoleus chthonoplastes TOW EF Microcoleus chthonoplastes CCY9605 GQ... Microcoleus chthonoplastes SAG 2210 E... Arthrospira sp. TJSD FJ Arthrospira jenneri EB 9604 GQ Arthrospira platensis EB 9602 GQ ZD 27F-1492R Geitlerinema cf. acuminatum CCALA Geitlerinema sp. CR 13M EF Şif Geitlerinema sp. PCC 7105 small subunit ribosomal RNA gene, partial sequence Length=1409 Score = 1578 bits (854), Expect = 0.0 Identities = 923/953 (96%), Gaps = 18/953 (1%) Strand=Plus/Plus Query 19 TGC-AGTCG-ACG-AGTCTTCGGACTTAGTGGCGGACGGGTGAGTAACGCGTGAGAACCT 75 Sbjct 29 TGCAAGTCGAACGAAGTCTTCGGACTTAGTGGCGGACGGGTGAGTAACGCGTGAGAACCT 88 Query 76 GCCTTGAGGAGGGGGATAACAGCGGGAAACTGCTGCTAATACCCCATATGCCGAAAGGTG 135 Sbjct 89 GCCTCGAGGAGGGGGATAACAGCGGGAAACTGCTGCTAATACCCCATATGCCGAAAGGTG 148 Query 136 AAAAGAAATTTGCCTTGAGAGGGGCTCGCGTCCGATTAGCTAGTTGGTGAGGTAAGAGCT 195 Sbjct 149 AAAAGAAATTTGCCTTGAGAGGGGCTCGCGTCCGATTAGCTAGTTGGTGAGGTAAGAGCT 208 Query 196 TACCAAGGCGACGATCGGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGAC 255 Sbjct 209 TACCAAGGCGACGATCGGTAGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGAC 268 Query 256 ACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTG 315
112 90 Sbjct 269 ACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTG 328 Query 316 ACGGAGCAACGCCGCGTGGGGGAAGAAGGCCTTTGGGTTGTAAACTCCTTTTCTCAGGGA 375 Sbjct 329 ACGGAGCAACGCCGCGTGGGGGAAGAAGGCCTTTGGGTTGTAAACTCCTTTTCTCAGGGA 388 Query 376 AGAAGAACTGACGGTACCTGAGGAATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGT 435 Sbjct 389 AGAAGAACTGACGGTACCTGAGGAATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGT 448 Query 436 AATACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTTCGTAGGCGGCGT 495 Sbjct 449 AATACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTTCGTAGGCGGCGT 508 Query 496 TTCAAGTCTGCTGTCAAAGGCCGAGGCTCAACTTCGGAAAGGCAGTGGAAACTGAAAAGC 555 Sbjct 509 TTCAAGTCTGCTGTCAAAGGCCGAGGCTCAACTTCGGAAAGGCAGTGGAAACTGAAAAGC 568 Query 556 TAGAGGTCGGTAGGGGCAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTGGG 615 Sbjct 569 TAGAGGTCGGTAGGGGCAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTGGG 628 Query 616 AAGAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGAACCTGACGCTGAGGGACGAAAGCT 675 Sbjct 629 AAGAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGAACCTGACGCTGAGGGACGAAAGCT 688 Query 676 AGGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCCGTAAACGATGGAGACTAGGT 735 Sbjct 689 AGGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCCGTAAACGATGGAGACTAGGT 748 Query 736 GTTGCCTGTATCGACCCGGGCAGTGCCGAAGCCAACGCGTTAAGTCTCCCGCCTGGGGAG 795 Sbjct 749 GTTGCCTGTATCGACCCGGGCAGTGCCGAAGCCAACGCGTTAAGTCTCCCGCCTGGGGAG 808 Query 796 TACGCACGCAAGTGTGAAACTCACAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGTAT 855 Sbjct 809 TACGCACGCAAGTGTGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGTAT 868 Query 856 GTGGATTTTATTTACGATGCAACGTCGTAGCATCTTTACCTAGGACTTGACCATCCCCGC 915 Sbjct 869 GTGG-TTTAATT--CGATGCAACG-CGAAGAACCTT-ACC-AGGGCTTGAC-ATCCC-GC 920 Query 916 GA-TCTCGGCGCAAGTAGATGAGTGACCTTACGTTA-CGCCGCAGACAAGGTG 966 Sbjct 921 GAATCTCGGCGAAAGTTGA-GAGTG-CCT-ACGGGAACGCGG-AGACA-GGTG 968 R Geitlerinema sp. PCC 7105 small subunit ribosomal RNA gene, partial sequence Length=1409 Score = 1452 bits (786), Expect = 0.0 Identities = 886/931 (95%), Gaps = 19/931 (2%) Strand=Plus/Minus Query 6 AGCCCCGCCTTCGG-ACCCTCCTCCGCGAACGGTTAGAGTAATGACTTCGGGCATGGCCG 64 Sbjct 1397 AGCCCTGCCTTCGGCACCCTCCTCCGCGAACGGTTAGAGTAATGACTTCGGGCATGGCCG 1338 Query 65 ACTTCCATGGTGTGACggggggggTGTACAAGGCCCGGGAACGGATTCACCGCACTATGC 124 Sbjct 1337 ACTTCCATGGTGTGACGGGCGGTGTGTACAAGGCCCGGGAACGGATTCACCGCAGTATGC 1278 Query 125 TGACCTGCGATTACTAGCGATTCCTCCTTCATGCAGGCGAGTTGCAACCTGCAATCTGAA 184 Sbjct 1277 TGACCTGCGATTACTAGCGATTCCTCCTTCATGCAGGCGAGTTGCAGCCTGCAATCTGAA 1218 Query 185 CTGAGGCCGGGTTTGAGAGATTAGCTCACTTTCGCAAGTTGGCTGCTCGCTGTCCCGACC 244 Sbjct 1217 CTGAGGCCGGGTTTGAGAGATTAGCTCACTTTCGCAAGTTGGCTGCTCGCTGTCCCGACC 1158 Query 245 ATTGTAGTACGTGTGTCGCCCAGGACATATGGGGCATGATGACTTGACGTCATCCCCACC 304 Sbjct 1157 ATTGTAGTACGTGTGTCGCCCAGGACATATGGGGCATGATGACTTGACGTCATCCCCACC 1098
113 91 Query 305 TTCCTCCGGTTTATCACCGGCAGTCTCCCTAGAGTGCCCCACTTAAGGATGGCAACTAAG 364 Sbjct 1097 TTCCTCCGGTTTATCACCGGCAGTCTCCCTAGAGTGCCCCACTTAAGGATGGCAACTAAG 1038 Query 365 GACGAGGGTTGCGCTCGTTGGGGGACTTAACCCAACATCTCACGACACGAGCTGACGACA 424 Sbjct 1037 GACGAGGGTTGCGCTCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACA 978 Query 425 GCCATGCACCACCTGTCTCCGCGTTCCCGTAGGCACTCTCAACTTTCGCCGAAATTCGCG 484 Sbjct 977 GCCATGCACCACCTGTCTCCGCGTTCCCGTAGGCACTCTCAACTTTCGCCGAGATTCGCG 918 Query 485 GGATGTCAAGCCCTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCG 544 Sbjct 917 GGATGTCAAGCCCTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCG 858 Query 545 CTTGTGCGGGCCCCCGTCAATTCCTTTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCG 604 Sbjct 857 CTTGTGCGGGCCCCCGTCAATTCCTTTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCG 798 Query 605 GGAGACTTGACGCGATGGCTTCGGCACTACGCGGGTCGATACAGGCAACACCTAGTCTCC 664 Sbjct 797 GGAGACTTAACGCGTTGGCTTCGGCACTGCCCGGGTCGATACAGGCAACACCTAGTCTCC 738 Query 665 ATCGTTTACGGATAGGACTACAGGGGTATCTATTCCCATTCGCTCCCCTAGCTTTCGTCC 724 Sbjct 737 ATCGTTTACGGCTAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCC 678 Query 725 CTCAACGTCAGGTTCGGTGCATCAGAGCGCTTTCGCCACCAGGTGTTCTTCCCAATATCC 784 Sbjct 677 CTCAGCGTCAGGTTCGGCCCAGCAGAGCGCTTTCGCCACC-GGTGTTCTTCCCAATATC- 620 Query 785 TACGCATTTCAACGCTACACTGGAGAAGTCCCATCTGCCCCGTACTGAC-TCTAGCCTTT 843 Sbjct 619 TACGCATTTCACCGCTACACTGG-GAATTCCC-TCTGCCCC-TACCGACCTCTAGC-TTT 564 Query 844 TCCAGATTCCACTGCCATTTCCGATGTATGACGCCTGCGGTCCTTTGAACAGCAGTACTG 903 Sbjct 563 TC-AGTTTCCACTGCC-TTTCCGAAGT-TGA-GCCT-CGG-CCTTTGA-CAGCAG-ACTT 512 Query 904 GAA-CTGCCACCTACGAACGACTTCACGCCC 933 Sbjct 511 GAAAC-GCCGCCTACGAACG-CTTTACGCCC Sif 27F-1492R 11 Geitlerinema sp. PCC 7105 AF Geitlerinema sp. PCC 7105 AB Geitlerinema sp. Flo1 FJ Geitlerinema sp. MBIC10006 AB Geitlerinema sp. A28DM FJ Geitlerinema sp. W-1 EF Geitlerinema sp. BBD P2b-1 EF Geitlerinema sp. BBD DQ Geitlerinema sp. BBD HS217 EF Geitlerinema sp. BBD HS223 DQ Geitlerinema sp. CCY0102 GQ Oscillatoriales cyanobacterium Schier Phormidiaceae cyanobacterium SAG
114 92 Bal ip Geitlerinema sp. PCC S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 and 23S rrna gene (partial), strain PCC 8501 Length=2024 Score = 1546 bits (837), Expect = 0.0 Identities = 879/898 (97%), Gaps = 8/898 (0%) Strand=Plus/Plus Query 16 CATGC-AGTCG-ACGGAGTCCTTCGGGACTTAGTGGCGGACGGGTGAGTAACGCGTGAGA 73 Sbjct 7 CATGCAAGTCGAACGGAGTGCTTCGGCACTTAGTGGCGGACGGGTGAGTAACGCGTGAGA 66 Query 74 ATCTGCCCTTAGGACGGGGACAACAGCTGGAAACGGCTGCTAATACCCGATGTGCCGAGA 133 Sbjct 67 ATCTGCCCTTAGGACGGGGACAACAGCTGGAAACGGCTGCTAATACCCGATGTGCCGAGA 126 Query 134 GGTGAAACATTTTATGGCCTGAGGATGAGCTCGCGTCTGATTAGCTAGTTGGTGGGGTAA 193 Sbjct 127 GGTGAAACATTTTATGGCCTGAGGATGAGCTCGCGTCTGATTAGCTAGTTGGTGGGGTAA 186 Query 194 GGGCCTACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGATCAGCCACACTGGGACT 253 Sbjct 187 GAGCCTACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGATCAGCCACACTGGGACT 246 Query 254 GAGACACGGCCCACACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGCAA 313 Sbjct 247 GAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGCAA 306 Query 314 GCCTGACGGAGCAAGACCGCGTGCGGGAGGACGGCCTATTGGTTGTAAACCGCTTTTGAT 373 Sbjct 307 GCCTGACGGAGCAAGACCGCGTGCGGGAGGACGGCCTATTGGTTGTAAACCGCTTTTGAT 366 Query 374 AGGGAAGAAGTTCTGACGGTACCTATCGAATCAGCATCGGCTAACTCCGTGCCAGCAGCC 433 Sbjct 367 AGGGAAGAAGTACTGACGGTACCTATCGAATCAGCATCGGCTAACTCCGTGCCAGCAGCC 426 Query 434 GCGGTAATACGGAGGATGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGT 493 Sbjct 427 GCGGTAATACGGAGGATGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGTAGGT 486 Query 494 GGTTGTTCAAGTCTGCTGTTAAAGCGTGCGGCTTAACCGCATACCAGCAGTGGAAACTGG 553 Sbjct 487 GGTTGTTCAAGTCTGCTGTTAAAGCGTGCGGCTTAACCGCATACCAGCAGTGGAAACTGG 546 Query 554 GCGACTAGAGTGCGGTAGGGGTCAGGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATA 613 Sbjct 547 GCGACTAGAGTGCGGTAGGGGTCAGGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATA 606 Query 614 TTGGGAAGAACACCGGTGGCGAAAGCGCCTGACTGGACCTGCACTGACACTGAGGGACGA 673 Sbjct 607 TTGGGAAGAACACCGGTGGCGAAAGCGCCTGACTGGACCTGCACTGACACTGAGGGACGA 666 Query 674 AAGCTAGGGGAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCCGTAAACGATGGACAC 733 Sbjct 667 AAGCTAGGGGAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCCGTAAACGATGGACAC 726 Query 734 TAGGTGTTGCCCGTATCGACCCGGGCAGTGCCGTATCCAACGCGTTAAGTGTCCCGCCTG 793 Sbjct 727 TAGGTGTTGCCCGTATCGACCCGGGCAGTGCCGTAGCCAACGCGTTAAGTGTCCCGCCTG 786 Query 794 GGGAGTACGCTCGCAAGCAGTGAAACTCAAAGGAATTGACGGGGGCCCCGCACAAGCG-T 852 Sbjct 787 GGGAGTACGCTCGCAAG-AGTGAAACTCAAAGGAATTGACGGGGGCCC-GCACAAGCGGT 844 Query 853 GCAGTATGTGATTAAATTCGATGCAACCGCGA-GAATCTTACCAGGCCTTCGACATGT 909 Sbjct 845 GGAGTATGTGGTTTAATTCGATGCAAC-GCGAAGAACCTTACCAGGGCTT-GACATGT 900
115 93 R Geitlerinema sp. PCC S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 and 23S rrna gene (partial), strain PCC 8501 Length=2024 Score = 1110 bits (601), Expect = 0.0 Identities = 639/656 (97%), Gaps = 8/656 (1%) Strand=Plus/Minus Query 16 CCTT-GGCGTCCTCATCCTTGCGGTTGGAGTAACGACTTCGGGCGTGGCCAACTCCCATG 74 Sbjct 1371 CCTTCGGCGTCCTCCTCCTTTCGGTTGGAGTAACGACTTCGGGCGTGGCCAACTCCCATG 1312 Query 75 GTGTGACGGGCGGTGTGTACAAGGCCCGGGAACGTATTCACCGCAGTATGCTGACCTGCG 134 Sbjct 1311 GTGTGACGGGCGGTGTGTACAAGGCCCGGGAACGTATTCACCGCAGTATGCTGACCTGCG 1252 Query 135 ATTACTAGCGATTCCTCCTTCATGCAGGCGAGTTGCAGCCTGCAATCTGAACTGAGCCAC 194 Sbjct 1251 ATTACTAGCGATTCCTCCTTCATGCAGGCGAGTTGCAGCCTGCAATCTGAACTGAGCCAC 1192 Query 195 GGTTTATGGGATTAGCTTGCTCTCGCGAGCTTGCAACCCTTTGTCCGCAGCATTGTAGTA 254 Sbjct 1191 GGTTTATGGGATTAGCTTGCTCTCGCGAGCTTGCAACCCTTTGTCCGCAGCATTGTAGTA 1132 Query 255 CGTGTGTAGCCCAGGACGTAAGGGGCATGCTGACTTGACGTCATCCCCACCTTCCTCCGG 314 Sbjct 1131 CGTGTGTAGCCCAGGACGTAAGGGGCATGCTGACTTGACGTCATCCCCACCTTCCTCCGG 1072 Query 315 TTTGTCACCGGCAGTCTCTCTAGAGTGCCCAACTGAATGATGGCAACTAAAAACGAGGGT 374 Sbjct 1071 TTTGTCACCGGCAGTCTCTCTAGAGTGCCCAACTGAATGATGGCAACTAAAAACGAGGGT 1012 Query 375 TGCGCTCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCATGCAC 434 Sbjct 1011 TGCGCTCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCATGCAC 952 Query 435 CACCTGTGCTCCAGTTCCCGAAGGCACCCCTCCCTTTCAGGAGGGTTCTGGACATGTCAA 494 Sbjct 951 CACCTGTGCTCCAGTTCCCGAAGGCACCCCTCCCTTTCAAGAGGGTTCTGGACATGTCAA 892 Query 495 GCCCTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTGCGG 554 Sbjct 891 GCCCTGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTGCGG 832 Query 555 GCCCCCGTCAATTTCGTTTGAGTTTCACTCTTGCGAGCGTACTCCCCAGGGCGAGACACT 614 Sbjct 831 GCCCCCGTCAATTCC-TTTGAGTTTCACTCTTGCGAGCGTACTCCCCAGG-CGGGACACT 774 Query 615 TAAACGCGTTGGACTACCGCACTGCCGCGGGATCGATATGGGCGAGCACCTACTGT 670 Sbjct 773 TAA-CGCGTTGG-CTACGGCACTGCC-CGGG-TCGATACGGGCAA-CACCTAGTGT 723
116 94 57 Leptolyngbya laminosa ETS-08 FM Oscillatoria sp. J-24-Osc AF Geitlerinema sp. PCC 8501 FM Cyanobacterium sp. (OS type I) L Filamentous thermophilic cyanobacteri Filamentous thermophilic cyanobacteri(2) Bal ip 27F-1492R KY Phormidium animale PMC partial 16S rrna gene, strain PMC Length=614 Score = 1090 bits (590), Expect = 0.0 Identities = 608/616 (98%), Gaps = 3/616 (0%) Strand=Plus/Plus Query 46 AGTGGCGGACGGGTGAGTAACGCGTGAGAATATAACAGT-AGGGGCGGGGATAACCCCTG 104 Sbjct 1 AGTGGCGGACGGGTGAGTAACGCGTGAGAATCT-ACCTTCA-GGTCGGGGATAACCCCTG 58 Query 105 GAAACGGGGGCTAATACCGGATAGGTCGAAAGATGAAAGGTTAACTGCCTGAAGAGGAGC 164 Sbjct 59 GAAACGGGGGCTAATACCGGATAGGTCGAAAGATGAAAGGTTAACTGCCTGAAGAGGGGC 118 Query 165 TCGCGTCTGATTAGCTAGTAGGTGTGGTAAGAGCGCACCTAGGCGACGATCAGTAGCTGG 224 Sbjct 119 TCGCGTCTGATTAGCTAGTAGGTGTGGTAAGAGCGCACCTAGGCGACGATCAGTAGCTGG 178 Query 225 TCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCA 284 Sbjct 179 TCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCA 238 Query 285 GCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACCGCGTGGGGGAGG 344 Sbjct 239 GCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAAGACCGCGTGGGGGAGG 298 Query 345 AAGGCCTTTGGGTTGTAAACCCCTTTTGTTAGGGAAGaaaaaaaTGACGGTACCTAACGA 404 Sbjct 299 AAGGCCTTTGGGTTGTAAACCCCTTTTGTTAGGGAAGAAAAAAATGACGGTACCTAACGA 358 Query 405 ATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTTATC 464 Sbjct 359 ATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTTATC 418 Query 465 CGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGCCCGGCAAGTCTGCTGTCAAAGACCAC 524 Sbjct 419 CGGAATTATTGGGCGTAAAGCGTCCGTAGGTGGCCCGGCAAGTCTGCTGTCAAAGACCAC 478 Query 525 AGCTCAACTGTGGAAAGGCAGTGGAAACTGCAAGGCTAGAGTGCGGTAGGGGTAGAGGGA 584 Sbjct 479 AGCTCAACTGTGGAAAGGCAGTGGAAACTGCAAGGCTAGAGTGCGGTAGGGGTAGAGGGA 538 Query 585 ATTCCCAGTGTAGCGGTGAAATGCGTAGAGATTGGGAAGAACACCGGTGGCGAAAGCGCT 644 Sbjct 539 ATTCCCAGTGTAGCGGTGAAATGCGTAGAGATTGGGAAGAACACCGGTGGCGAAAGCGCT 598 Query 645 CTACTGGGCCGCAACT 660 Sbjct 599 CTACTGGGCCGCAACT 614 R Phormidium cf. terebriformis AB2002/07 16S ribosomal RNA gene, partial sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; 23S ribosomal RNA gene, partial sequence;
117 95 and trna-ile and trna-ala genes, complete sequence Length=1872 Score = 1182 bits (640), Expect = 0.0 Identities = 775/840 (92%), Gaps = 10/840 (1%) Strand=Plus/Minus Query 27 TCCTCATCCGCGAACGGTTGGAGTAACGACTTCGGGCGTGGCCAGCTCCCATGGTGTGAC 86 Sbjct 1325 TCCTCCTCCGCGAACGGTTAGAGTAACGACTTCGGGCGTGGCTAACTTCCATGGTGTGAC 1266 Query 87 GGGCGGTGTGTACAAGGCCCGGGAACGGATTCACCGCCGTATGCTGACCGGCGATTACTA 146 Sbjct 1265 GGGCGGTGTGTACAAGGCCCGGGAACGGATTCACCGCAGTATGCTGACCTGCGATTACTA 1206 Query 147 GCGATTCCGCCTTCACGCAGGCGAGTTGCAACCTGCGATCTGAACTGAGCCGAGGTTTAT 206 Sbjct 1205 GCGATTCCTCCTTCATGCAGGCGAGTTGCAGCCTGCAATCTGAACTGAGGAGAGGTTTAC 1146 Query 207 GAGATTAGCTC-GCCCTCGCGGGTTGGCTGCTCTTTGTCCACAGCATTGTATTACGTGTG 265 Sbjct 1145 GGGATTGGCTCAG-ACTCGCGTCTTGGCTACCCTTTGTCCCCCCCATTGTAGTACGTGTG 1087 Query 266 TCGCCCAGGACGTAAGGGGCATGCTGACTTGACGTCATCCCCACCTTCCTCCGGTTTGTC 325 Sbjct 1086 TAGCCCAGGACGTAAGGGGCATGATGACTTGACGTCATCCCCACCTTCCTCCGGTTTGTC 1027 Query 326 ACCGGCAGTCTCCCTAGAGTGCCCATCCAAAATGCTGGCAACTAAGGACGAGGGTTGCGC 385 Sbjct 1026 ACCGGCAGTCTCCCTAAAGTGCCCA-ACTAAATGATGGCAACTAAGGACGAGGGTTGCGC 968 Query 386 TCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCATGCACCACCT 445 Sbjct 967 TCGTTGCGGGACTTAACCCAACATCTCACGACACGAGCTGACGACAGCCATGCACCACCT 908 Query 446 GTGTTCCGGCTCCCGAAGGCACCCTCTGCTTTCA-CAGAGGTTCCGGACATGTCAAGCCC 504 Sbjct 907 GTGTTCCGGCTCCCGAAGGCACTCTCGGCTTTCACCA-AGATTCCGGACATGTCAAGCCC 849 Query 505 TGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTGCGGGCCC 564 Sbjct 848 TGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTGCGGGCCC 789 Query 565 CCGTCAATTCCTTTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCGGGAAACTTAACGC 624 Sbjct 788 CCGTCAATTCCTTTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCGGGAAACTTAACGC 729 Query 625 GATAGCTCCGATACAGCACGGGTCGATAC-GTGCTACACCTAGTTTACATCGTATACAGC 683 Sbjct 728 GTTAGCTTCGGCACAGCCCGGGTCGATACAG-GCTACACCTAGTTTCCATCGTTTACGGC 670 Query 684 TAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCCATCAATGACAGT 743 Sbjct 669 TAGGACTACTGGGGTATCTAATCCCATTCGCTCCCCTAGCTTTCGTCCCTCAGTGTCAGT 610 Query 744 TGCGGCCCATTAGAGCGCTTTCGCCACCAGTGTTCTTCCCAATCTCTACGCATTTCACCG 803 Sbjct 609 TGCGGCCCAGCAGAGCGCTTTCGCCACCGGTGTTCTTCCCGATATCTACGCATTTCACCG 550 Query 804 CTACACTGGGAATTGCACTCTACCCCTACCGCACTCTAGCCCTTGCCAGTTTCCACTGCC 863 Sbjct 549 CTACACCGGGAATTCC-CTCTGCCCCTACCGCACTCTAGCTGTT-C-AGTTTCCACTGCC 493
118 96 92 Lyngbya birgei CCC 333 EU Phormidium animale NTMP03 GU Phormidium animale PMC AJ KY 27F-1492R Microcoleus chthonoplastes CCY0602 GQ... Oscillatoria spongeliae 310P1 AY Oscillatoria spongeliae 32P1 AY Oscillatoria spongeliae 31P1 AY U sal Halospirulina sp.'ccc Baja-95 Cl.3', 16S rrna gene Length=1444 Score = 1395 bits (755), Expect = 0.0 Identities = 882/939 (93%), Gaps = 28/939 (2%) Strand=Plus/Plus Query 18 CATGC-AGTCG-ACGG-TCTCTTAGGAGAT-AGTGGCGGACGGGTGAGTAACGCGTGAGA 73 Sbjct 31 CATGCAAGTCGAACGGAT-TCTTAGGA-ATCAGTGGCGGACGGGTGAGTAACACGTGAAA 88 Query 74 ACCTGCCTCTAGGACGGGGACAACAGAGGGAAACTTCTGCTAATTCCCGAT-ACGCCTTT 132 Sbjct 89 ACCTGCCTCTAGGACGGGGATAACAGAGGGAAACTTCTGCTAATTCCCGATGA-GCCTGC 147 Query 133 GGGTAAAA-GATTTATCGCCTAGAGAGGGGCTCGCGTCCGATTAGCTAGTTGGTGGGGTA 191 Sbjct 148 GGGTAAAAGGATTTATCGCCTAGAGAGGGGTTCGCGTCCGATTAGCTAGTTGGTGGGGT- 206 Query 192 AAAGCCT-ACCAAGGCAGCGATCGGTAGCTGTTCTGAGAGGAAGAACAGCCACACTGGGA 250 Sbjct 207 AAAGGCTCACCAAGGCTGCGATCGGTAGCTGTTCTGAKAGGAAGAACAGCCACACTGGGA 266 Query 251 CTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGC 310 Sbjct 267 CTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGA 326 Query 311 AAGCCTGACGGAGCAATACCGCGTGAGGGAAGAAGGTCTTTGGATCGTAAACCTCTTTTC 370 Sbjct 327 AAGCCTGACGGAGCAATACCGCGTGAGGGAAGAAGGTCTTTGGATCGTAAACCTCTTTTC 386 Query 371 TCAGGGAAGAAGATCTGACGGTACCTGAGGAATAAGCCTCGGCTAACTCCGTGCCAGCAG 430 Sbjct 387 TCAGGGAAGAAGTTCTGACGGTACCTGAGGAATAAGCCTCGGCTAACTCCGTGCCAGCAG 446 Query 431 CCGCGGTAATACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTTCGCAG 490 Sbjct 447 CCGCGGTAATACGGAGGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGTTCGCAG 506 Query 491 GTGGCTATTCAAGTCTGCTGTCAAAGACCAGAGCTCAACTCTGGGTAGGCGGTGGAAACT 550 Sbjct 507 GTGGCTCGTCAAGTCTGCTGTCAAAGACCGGAGCTTAACTCCGGGTAGGCGGTGGAAACT 566 Query 551 GAATAGCTAGAGTGCGGTAGGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGA 610 Sbjct 567 GAAGAGCTAGAGTGCGGTAGGGGTAGAGGGAATTCCCAGTGTAGCGGTGAAATGCGTAGA 626 Query 611 GATTGGGAAGAACATCGGTGGCGAAAGCGCTCTACT-GGACCGCAACTGACACTCAGGGA 669 Sbjct 627 GATTGGGAAGAACACCGGTGGCGAAAGCGCTCTACTAGG-CCGCAACTGACACTCAGGGA 685 Query 670 CGAAAGCTAGGGGAGCGAAAGGGATTAGATACCCCTGTAGTCCTAGCCGTAAACGATGGG 729 Sbjct 686 CGAAAGCTAGGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCTGTAAACGATGGG 745
119 97 Query 730 TACTAGGCGTAGATTGTATCAACTCAAGCTGTGCCGAAGCTAACGCGTTAAGTACCCCGC 789 Sbjct 746 TACTAGGCGTAGCTTGTATCGACCCGAGCTGTGCCGAAGCTAACGCGTTAAGTACCCCGC 805 Query 790 CTGGGGAGTACGCACGCAAGTGTGAAACTCAAAGGAATTGACGGGGGGCCCGCACAAGCG 849 Sbjct 806 CTGGGGAGTACGCACGCAAGTGTGAAACTCAAAGGAATTGACGGGGG-CCCGCACAAGCG 864 Query 850 GTGGAGTATGTGGGTTTAATTTCGATGCCACCGCCGA-GAACCCTTACCCAGGGCTTGGA 908 Sbjct 865 GTGGAGTATGTGG-TTTAATT-CGATGCAAC-GC-GAAGAACC-TTACC-AGGGCTTG-A 917 Query 909 CCATCCTGGCCGAAATTTTTCTGGCAAACAGAGTAGTGC 947 Sbjct 918 C-ATCC-GGT-GAA-TTCT-CTGG-AAACAGAGAAGTGC 950 R Halospirulina sp.'ccc Baja-95 Cl.3', 16S rrna gene Length=1444 Score = 1136 bits (615), Expect = 0.0 Identities = 761/828 (91%), Gaps = 24/828 (2%) Strand=Plus/Minus Query 37 GTTAGAGTAACGACTTCGGATGTTGCCAGCTTCCATGGTGTGACGGGCGGTGGGTACAAG 96 Sbjct 1371 GTTGGAGTAACGACTTCGGATGTTGCCAGCTTCCATGGTGTGACGGGCGGTGTGTACAAG 1312 Query 97 GCCCGGAAACGGATTCACCGTAC-TATTCTGACCTGG-GATTACTAACGATTCCTCCTTC 154 Sbjct 1311 GCCCGGGAACGAATTCACCG-CCGTATGCTGACC-GGCGATTACTAGCGATTCCTCCTTC 1254 Query 155 ACGCAGGTGAGTTGCAACCTGCGATCTGAACTGAGGCTGAGTTTGATGGGATTCGCTGGT 214 Sbjct 1253 ACGCAGGCGAGTTGCAGCCTGCGATCTGAACTGAGGCCGAGTTTGATGGGATTCGCT--A 1196 Query 215 CCTCTCG-GA--TTCGCTGCCCTTTGTCTCAACCATTGTA-AACGTGTGTAGCCCAGGAC 270 Sbjct 1195 ACTCTCGCGAGCTT-GCTGCCCTTTGTCTCGACCATTGTAGTACGTGTGTTGCCCAGGAC 1137 Query 271 GTAAGGGGCATGCTGACTTGACTTCATCCCCGCCTTCCTCCGGTTTGTCACCGGAAGTCT 330 Sbjct 1136 GTAAGGGGCATGCTGACTTGACGTCATCCCCACCTTCCTCCGGTTTGTCACCGGCAGTCT 1077 Query 331 CCCTAGAGTGCCCAACTTAATGATGGCAACTAAGGACGAGGGTTGCGCTCGTTGCGGGAC 390 Sbjct 1076 CCCTAAAGTGCCCAACTTAATGCTGGCAACTAAGGACGAGGGTTGCGCTCGTTGCGGGAC 1017 Query 391 TTAACCCAACATCTCACGACACGAGCTGACGACAGCCATGCACCACCTGTCACGGCGCTC 450 Sbjct 1016 TTAACCCAACATCTCACGACACGAGCTGACGACAGCCATGCACCACCTGTCACGGTGCTC 957 Query 451 CC-TAAGGCACTTCTCTGTTTCCAGAAAATTCGCCGGATGTCAAGCCCTGGTAAGGTTCT 509 Sbjct 956 CCGTA-GGCACTTCTCTGTTTCCAGAGAATTCACCGGATGTCAAGCCCTGGTAAGGTTCT 898 Query 510 TCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTGCGGGCCCCCGGCAATTCCT 569 Sbjct 897 TCGCGTTGCATCGAATTAAACCACATACTCCACCGCTTGTGCGGGCCCCCGTCAATTCCT 838 Query 570 TTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCGGGATACTTAACGCGTTAGCTTCGGC 629 Sbjct 837 TTGAGTTTCACACTTGCGTGCGTACTCCCCAGGCGGGGTACTTAACGCGTTAGCTTCGGC 778 Query 630 ACAGCTTGAGTTGATACAATCTACGCCTAGTACCCATCGATTACGGCTAGGACTACAGGG 689 Sbjct 777 ACAGCTCGGGTCGATACAAGCTACGCCTAGTACCCATCGTTTACAGCTAGGACTACTGGG 718 Query 690 GTATCAAATCCCTTTCGCTCCCCTAGCTATCGATCCCTGAGTGTCAGTTGCGGTCC-AGT 748 Sbjct 717 GTATCTAATCCCATTCGCTCCCCTAGCTTTCG-TCCCTGAGTGTCAGTTGCGG-CCTAGT 660 Query 749 AGAGCGCTGTCGCCACCGATGTTCTTCCCAATCTCTACGCTGTTTCAGCCGCTACGCTGG 808
120 98 Sbjct 659 AGAGCGCTTTCGCCACCGGTGTTCTTCCCAATCTCTACGCA-TTTCA-CCGCTACACTGG 602 Query 809 GGATTACACTCTACCCCATATCGACAGCTCTAGCCTATTCAGTTGTCC 856 Sbjct 601 GAATTCC-CTCTACCCC-TACCG-CA-CTCTAGC-TCTTCAGTT-TCC Spirulina subsalsa FACHB351 FJ Spirulina subsalsa FACHB351 AF Spirulina subsalsa AB2002/06 AY Spirulina subsalsa IAM M-223 AB Spirulina sp. MPI S4 Y Spirulina sp. P7 AF U sal 27F-1492R Halospirulina tapeticola CCC Baja Halospirulina sp. CCC Baja-95 Cl.3 Y1... Halospirulina sp. MPI S3 Y Microcoleus chthonoplastes WW6 EF Microcoleus chthonoplastes WW5 EF U ip Geitlerinema sp. CCY S ribosomal RNA gene, partial sequence Length=1299 Score = 1221 bits (661), Expect = 0.0 Identities = 809/877 (92%), Gaps = 23/877 (2%) Strand=Plus/Plus Query 45 AGTGGCGGACGGGTGAGTAACGCGTGAGAATCTGCCTTCAGGACGGGGACAACAGCTGGA 104 Sbjct 1 AGTGGCGGACGGGTGAGTAACGCGTGAGAATCTACCTTCAGGACGGGGACAACAGCTGGA 60 Query 105 AACGGCTGCTAATACCCGATGTGCCGAGAGGTGAAATGTT-TTATCGCCTGAAGAGGAGC 163 Sbjct 61 AACGGCTGCTAATACCCGATATGCCGAAAGGTGAAAAGTTATT-TCGCCTGGAGAGGAGC 119 Query 164 TCGCGTCCGATTAGCTAGTTGGCGGGGTAAAAGCCCACCAAAGCGACGATCGGTAGCTGG 223 Sbjct 120 TCGCGTCCGATTAGCTAGTTGGTGGGGTAAAAGCTTACCAAGGCGACGATCGGTAGCTGG 179 Query 224 TCTGAGAGGACGAGCAGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCA 283 Sbjct 180 TCTGAGAGGATGAGCAGCCACACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCA 239 Query 284 GCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAACGCCGCGTGGGGGAAG 343 Sbjct 240 GCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGGAGCAACGCCGCGTGGGGGAAG 299 Query 344 ACTGCCCTTGGGTTGTAAACCCCTTTTCTCAGGGAAG-AC-CCCCGACGGTACCTGAGGA 401 Sbjct 300 AAGGTTTTTGGATTGTAAACCCCTTTTCTCAAGGAAGAACAAAATGACGGTACTTGAGGA 359 Query 402 AGAAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTTATC 461 Sbjct 360 ATCAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTTATC 419 Query 462 CGGAATTATTGGGCGTAAAGCGTTCGTAGGCGGC-GTATCAAGTCTGCGGTCAAA-ATCC 519 Sbjct 420 CGGAATTATTGGGCGTAAAGCGTTCGTAGGCGGCTGT-TCAAGTCTGCTGTCAAAGA-CC 477 Query 520 GAGGCTCAACCTCGGGCCGGCCGTGGAAACTGAA-ACGCTAGAG-TACGGTAGGGGTAGA 577
121 99 Sbjct 478 GAGGCTCAACCTCGGGTCGGCAGTGGAAACTGAATA-GCTAGAGGT-CGGTAGGGGCAGA 535 Query 578 GGGAATTCCCGGTGTAGCGGTGAAATGCGTAGATATCGGGAAGAACACCGGTGGCGAAAG 637 Sbjct 536 GGGAATTCCCAGTGTAGCGGTGAAATGCGTAGATATTGGGAAGAACACCGGTGGCGAAAG 595 Query 638 CGCTCTACTGGGCCGAAACTGACGCTGAGGAACGAAAGCTAGGGGAGCGAATGGGATTAG 697 Sbjct 596 CGCTCTGCTGGGCCGAACCTGACGCTGAGGGACGAAAGCTAGGGGAGCGAATGGGATTAG 655 Query 698 ATACCCCAGTAGTCCTAGCCGTAAACGATGGAGACTAGGTGTAGTCC-GTATCGACCCGG 756 Sbjct 656 ATACCCCAGTAGTCCTAGCCGTAAACGATGGAAACTAGGTGTGG-CCTGTATCGACCCGG 714 Query 757 ACTGTGCCGAAGCCAACGCGTTAAGTCTCCCGCCTGGGGAGTACGCGCGCAAGCGTGAAA 816 Sbjct 715 GCCGTGCCGAAGCTAACGCGTTAAGTTTCCCGCCTGGGGAGTACGCACGCAAGTGTGAAA 774 Query 817 CTCAAAGGGAATTGACGGGGGCCCGCACAAGCGGTGGGAGTATGTGGTTTAATTTCGATG 876 Sbjct 775 CTCAAAGG-AATTGACGGGGGCCCGCACAAGCGGTGG-AGTATGTGGTTTAATT-CGATG 831 Query 877 CCAACCGCGA-GAACCTTTACCCAGGACTGGACCATG 912 Sbjct 832 C-AAC-GCGAAGAACCTT-ACC-AGGGCTTGAC-ATG 863 R Geitlerinema sp. CCY S ribosomal RNA gene, partial sequence Length=1299 Score = 1230 bits (666), Expect = 0.0 Identities = 848/931 (91%), Gaps = 31/931 (3%) Strand=Plus/Minus Query 41 CTTCGGGCGTGGCCGGCTTCCATGGTGTGACGGGCGGTGTGTACAAGGCCCGGGAACGGA 100 Sbjct 1299 CTTCGGGCATGGCCAACTTCCATGGTGTGACGGGCGGTGTGTACAAGGCCCGGGAACGGA 1240 Query 101 TTCACCGCGGTATGCTGACCCGCGATTACTAGCGATTCCTCCTTCAAGCAGGCGAGTTGC 160 Sbjct 1239 TTCACCGCAGTATGCTGACCTGCGATTACTAGCGATTCCTCCTTCATGCAGGCGAGTTGC 1180 Query 161 AGCCTGCTATCCGAACTGGGGCCGGGTTTAAGGGATTAGCTCGCTCTTGCGAGTTGGCTG 220 Sbjct 1179 AGCCTGCAATCTGAACTGAGGCCGGGTTTAAGAGATTAGCGCACTCTCGCGAGTTGGCTG 1120 Query 221 CCCGTTGTCCCGACCATTGTAGTACGTGTGTCGCCCAGGGCATAAGGGGCATGCTGACTT 280 Sbjct 1119 CTCGTTGTCCCGACCATTGTAGTACGTGTGTAGCCCAGGACATATGGGGCATGATGACTT 1060 Query 281 GACGTCATCCCCACCTTCCTCCGGTTTATCACCGGCAGTCTCTCTAGAGTGCCCATCCAA 340 Sbjct 1059 GACGTCATCCCCACCTTCCTCCGGTTTATCACCGGCAGTCTCCCTAGAGTGCCCA-ACTT 1001 Query 341 AATGCTGGCAACTAAAGACGAGGGTTGCGCTCGTTGCGGGACTTAACCCAACATCTCACG 400 Sbjct 1000 AATGATGGCAACTAAGGACGAGGGTTGCGCTCGTTGCGGGACTTAACCCAACATCTCACG 941 Query 401 ACACGAGCTGACGACAGCCATGCACCACCTGTGTTCCG-GCTCCCGTAGGCACTCCTC-C 458 Sbjct 940 ACACGAGCTGACGACAGCCATGCACCACCTGTG-TCCGCGCTCCCGTAGGCACT-CTCTC 883 Query 459 CTTTCAGGCG-GATTC-CGGACATGTCAAGCCCTGGTAAGGTTCTTCGCGTTGCATCGAA 516 Sbjct 882 CTTTCA-GAGAGATTCGCGG-CATGTCAAGCCCTGGTAAGGTTCTTCGCGTTGCATCGAA 825 Query 517 TTAAACCACATACTCCACCGCTTGTGCGGGCCCCCGTCAATTCCTTTGAGTTTCACGCTT 576 Sbjct 824 TTAAACCACATACTCCACCGCTTGTGCGGGCCCCCGTCAATTCCTTTGAGTTTCACACTT 765
122 100 Query 577 GCGCGCGTACTCCCCAGGCGGGAGACTTAACGCGTTGGCTTCGGCACAGTCCGGGTCGAT 636 Sbjct 764 GCGTGCGTACTCCCCAGGCGGGAAACTTAACGCGTTAGCTTCGGCACGGCCCGGGTCGAT 705 Query 637 AC-GGACTACACCTAGTCTCCATCGTTTACGGCTAGGACTACTGGGGTATCTAATCCCAT 695 Sbjct 704 ACAGGCC-ACACCTAGTTTCCATCGTTTACGGCTAGGACTACTGGGGTATCTAATCCCAT 646 Query 696 TCGCTCCCCTAGCTTTCGTTCCTCAGCGTCAGTTTCGGCCCAGTAGAGCGCTTTCGCCAC 755 Sbjct 645 TCGCTCCCCTAGCTTTCGTCCCTCAGCGTCAGGTTCGGCCCAGCAGAGCGCTTTCGCCAC 586 Query 756 CGGTGTTCTTCCCGATATCTACGCATTTCACCGCTACACCGGGAAATTCCCTCTACCCCT 815 Sbjct 585 CGGTGTTCTTCCCAATATCTACGCATTTCACCGCTACACTGGGAA-TTCCCTCTGCCCCT 527 Query 816 ACCGTAC-TCTAGCGT-TTCAGTTTCCACGGGCCGGCCCGAGGTTTGAGCCCTCGGGATT 873 Sbjct 526 ACCG-ACCTCTAGC-TATTCAGTTTCCACTG-CCGACCCGAGGTT-GAGCC-TCGGTCTT 472 Query 874 TTGACCGACAGACTTTGATAC-GCTCGACTTACGGACGGCTTTACGCCCAAATA-TGCAG 931 Sbjct 471 T-GACAG-CAGACTT-GA-ACAGC-CGCCT-ACGAACG-CTTTACGCCCAA-TAATTCCG 420 Query 932 -ATTACGCTTGACTCGTACGCTATTACCGCG 961 Sbjct 419 GATAACGCTTGCCTCCTCCG-TATTACCGCG 390 Oscillatoriales cyanobacterium Schier Phormidiaceae cyanobacterium SAG Phormidium sp. HBC9 EU Geitlerinema sp. CCY0102 GQ U sal 27F-1492R 0.01 Çizelge 4.3: 16SF-23SR Dizi analizi sonuçları Primerler Organizma Adı Maksimum 1 BF (16SF- 23SR) U pse Oscillatoriales cyanobacterium JSC-1 16S-23S ribosomal RNA intergenic spacer, partial sequence 1 BR (16SF-23SR) U pse Oscillatoriales cyanobacterium JSC-1 16S-23S ribosomal RNA % 82 intergenic spacer, partial sequence 2 BF (16SF- 23SR) U osc Oscillatoria cf. curviceps Fkv-4 16S-23S ribosomal RNA intergenic % 85 spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence 2 BR (16SF-23SR) U osc Oscillatoria cf. curviceps Fkv-4 16S-23S ribosomal RNA intergenic % 93 spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence 3 BF (16SF- 23SR) KK Phormidium autumnale UTCC S rrna gene (partial), 23S %95 rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 3 BR (16SF- 23SR) KK Phormidium autumnale UTCC S rrna gene (partial), 23S % 95 rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 4 BF (16SF-23SR) Bal pse Uncultured cyanobacterium isolate DGGE gel band 3 16S ribosomal % 96 RNA gene, partial sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; and 23S ribosomal RNA gene, partial sequence 4 BR(16SF-23SR) Bal pse Uncultured cyanobacterium isolate DGGE gel band 3 16S ribosomal % 94 RNA gene, partial sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; and 23S ribosomal RNA gene, partial sequence 5 BF (16SF- 23SR) ZD Geitlerinema cf. acuminatum CCALA S-23S ribosomal RNA % 85 intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence 5 BR (16SF- 23SR) ZD Geitlerinema cf. acuminatum CCALA S-23S ribosomal RNA % 81 intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence 6 BF (16SF- 23SR) Şif Geitlerinema sp. PCC S-23S ribosomal RNA intergenic % 99 Tanımlanma % 83
123 101 spacer, complete sequence 6 BR (16SF- 23SR) Şif Geitlerinema sp. PCC S-23S ribosomal RNA intergenic spacer, complete sequence 7 BF (16SF- 23SR) Bal ip Leptolyngbya frigida ANT.LH S-23S ribosomal RNA intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence 7 BR (16SF- 23SR) Bal ip Leptolyngbya frigida ANT.LH S-23S ribosomal RNA intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence 8 BF (16SF- 23SR) KY Phormidium autumnale UTCC S rrna gene (partial), 23S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 8 BR(16SF- 23SR) KY Phormidium autumnale UTCC S rrna gene (partial), 23S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 9 BF (16SF- 23SR)U sal Uncultured marine bacterium clone SPOTSAPR01_14m146 16S ribosomal RNA gene, partial sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; and 23S ribosomal RNA gene, partial sequence 9 BR (16SF- 23SR) U sal Uncultured marine bacterium clone SPOTSAPR01_14m146 16S ribosomal RNA gene, partial sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; and 23S ribosomal RNA gene, partial sequence 10 BF (16SF- 23SR)U ip Phormidium persicinum SAG S rrna gene (partial), ITS1, 23S rrna gene (partial), trna-ile gene and trna-ala gene, strain SAG BR (16SF- 23SR)U ip Phormidium persicinum SAG S rrna gene (partial), ITS1, 23S rrna gene (partial), trna-ile gene and trna-ala gene, strain SAG % 98 % 93 % 93 % 95 % 95 % 83 % 83 %95 %95 U pse 16SF-23SR Oscillatoriales cyanobacterium JSC-1 16S-23S ribosomal RNA intergenic spacer, partial sequence Length=449 Score = 204 bits (110), Expect = 6e-49 Identities = 211/254 (83%), Gaps = 29/254 (11%) Strand=Plus/Plus Query 84 CTTTCAAA-T-TTAGGTTCGTGGAATGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCT 141 Sbjct 90 CTTTCAAACTATTTGGTTCGAGGAATGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCT 149 Query 142 GATAAGGGTGAGGTCCCTGGTTCAAGTCCAGGATGGCCCAC-TTATCAGTAAGC-A-GA- 197 Sbjct 150 GATAAGGGTGAGGTCCCTGGTTCGAGTCCAGGATGGCCCACCTTAACAGGA-GCGATGAG 208 Query 198 TATCAGATATCAGATTTCAAGAGATTGATAG-CTGGTGGCTGGTGGCTGATAACTGAAAT 256 Sbjct 209 TGTCGGAGGT-AGATT-C--GAGATTCA-AGACT-----CT--TGG--GATA--TG Query 257 GGGGGTATAGCTCAGTTGGTAGAGCGCCTGCTTTGCAAGCAGGATGTCAGCGGTTCGAGT 316 Sbjct 249 GGGGTT-TAGCTCAGTTGGTAGAGCGCCTGCTTTGCAAGCAGGATGTCAGCAGTTCGAGT 307 Query 317 CCGCTTACCTCCAC 330 Sbjct 308 CTGCTAACCTCCAC 321 R Oscillatoriales cyanobacterium JSC-1 16S-23S ribosomal RNA intergenic spacer, partial sequence Length=449 Score = 200 bits (108), Expect = 7e-48 Identities = 209/252 (82%), Gaps = 29/252 (11%) Strand=Plus/Minus
124 102 Query 205 GGAGGTAAGCGGACTCGAACCGCTGACATCCTGCTTGCAAAGCAGGCGCTCTACCAACTG 264 Sbjct 319 GGAGGTTAGCAGACTCGAACTGCTGACATCCTGCTTGCAAAGCAGGCGCTCTACCAACTG 260 Query 265 AGCTATACCCCCATTTCAGTTATCAGCCACCAGCCACCAG-CTATCAATCTCTTGAAATC 323 Sbjct 259 AGCTA-AACCCC----CA--TATC--CCA--AG-----AGTCT-TGAATCTC--G-AATC 220 Query 324 TGATATCTGATA-TC-T-GCTTACTGATAA-GTGGGCCATCCTGGACTTGAACCAGGGAC 379 Sbjct 219 T-ACCTCCGACACTCATCGC-TCCTGTTAAGGTGGGCCATCCTGGACTCGAACCAGGGAC 162 Query 380 CTCACCCTTATCAGGGGTGCGCTCTAACCACCTGAGCTAATAGCCCATTCCACGAACCTA 439 Sbjct 161 CTCACCCTTATCAGGGGTGCGCTCTAACCACCTGAGCTAATAGCCCATTCCTCGAACCAA 102 Query 440 A-A-TTTGAAAG 449 Sbjct 101 ATAGTTTGAAAG Phormidium sp. OL 81 AM Phormidium sp. OL S4 AM U Pse 16S-23S Arthrospira sp. PD2003/03 FJ Arthrospira sp. PD2002/08 FJ Scytonema hyalinum FI-8A AF Leptolyngbya valderiana BDU AJ... Leptolyngbya valderiana BDU AJ Geitlerinema sp. Flo1 FJ Geitlerinema sp. PCC 7105 FJ U osc Oscillatoria cf. curviceps Fkv-4 16S-23S ribosomal RNA intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence Length=449 Score = 152 bits (82), Expect = 1e-33 Identities = 135/158 (85%), Gaps = 13/158 (8%) Strand=Plus/Plus Query 10 AGCTTGGAGATCCCAGGTCGTTCGAGGTCAAC-TCAGaaa--aaaaGTGATAAAGCTTTC 66 Sbjct 49 AGC-TGGACATCCCTGGTCGTACAAGG-GAACATCA-AAAGTCAAAGT--T-AA-CTTTC 101 Query 67 AAAC--GAATAGGGTTCGGTT-AGGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGA 123 Sbjct 102 AAACTAGAGTCTGGTTCGGTTCATGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGA 161 Query 124 TAAGGGTGAGGTCCCTGGTTCAAGTCCAGGATGGCCCA 161 Sbjct 162 TAAGGGTGAGGTCCCTGGTTCAAGTCCAGGATGGCCCA 199 R Oscillatoria cf. curviceps Fkv-4 16S-23S ribosomal RNA intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence Length=449
125 103 Score = 148 bits (80), Expect = 2e-32 Identities = 96/103 (93%), Gaps = 3/103 (2%) Strand=Plus/Minus Query 215 TGGGCCATCCTGGACTTGAACCAGGGACCTCACCCTTATCAGGGGTGCGCTCTAACCACC 274 Sbjct 199 TGGGCCATCCTGGACTTGAACCAGGGACCTCACCCTTATCAGGGGTGCGCTCTAACCACC 140 Query 275 TGAGCTAATAGCCCCT-AACCGAACCCTATTC--GTTTGAAAG 314 Sbjct 139 TGAGCTAATAGCCCATGAACCGAACCAGACTCTAGTTTGAAAG Arthrospira sp. PD2003/03 FJ Arthrospira sp. PD2002/08 FJ A. jenneri EB 9604 GQ A. erdosensis EB 9601 GQ Arthrospira maxima BJ 2000 GQ A. platensis NJ 1999 GQ Blennothrix sp. AQS EU U osc 16S-23S 0.05 KK Phormidium autumnale UTCC S rrna gene (partial), 23S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 Length=593 alignments for this subject sequence by: Score Percent identity start position Subject start position Score = 172 bits (93), Expect = 2e-39 Identities = 109/116 (93%), Gaps = 4/116 (3%) Strand=Plus/Plus Sort E value Query Query 78 TTTCAAAC--TGAGTTCGGTTCGGAGTAAGGGCTATTAGCTCAGGTGGTTAGAGCGCACC 135 Sbjct 139 TTTCAAACTATCAATT-GGTTCGGAGTATGGGCTATTAGCTCAGGTGGTTAGAGCGCACC 197 Query 136 CCTGATAAGGGTGAGGTCCCTGGTTCAAGTCCAGGATGGCCCACCTGAACCCCAAA 191 Sbjct 198 CCTGATAAGGGTGAGGTCCCTGGTTCAAGTCCAGGATGGCCCACCT-AACCCCAAA 252 Score = 124 bits (67), Expect = 5e-25 Identities = 77/81 (95%), Gaps = 4/81 (4%) Strand=Plus/Plus Query 267 GGGGGTATAGCTCAGTTGGTAGAGCGCCTG-CTTTGCAA-GCAGGATGTCAGCGGTTCGA 324 Sbjct 257 GGGGGTATAGCTCAGTTGGTAGAGCG-CTGCCTTTGCAAGGCA-GATGTCAGCGGTTCGA 314 Query 325 GTCCGCTTACCTCCACCAAAA 345 Sbjct 315 GTCCGCTTACCTCCACCAAAA 335
126 104 R Phormidium autumnale UTCC S rrna gene (partial), 23S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 Length=593 alignments for this subject sequence by: Score Percent identity start position Subject start position Score = 172 bits (93), Expect = 2e-39 Identities = 109/116 (93%), Gaps = 4/116 (3%) Strand=Plus/Minus Sort E value Query Query 351 TTTGGGGTTCAGGTGGGCCATCCTGGACTTGAACCAGGGACCTCACCCTTATCAGGGGTG 410 Sbjct 252 TTTGGGGTT-AGGTGGGCCATCCTGGACTTGAACCAGGGACCTCACCCTTATCAGGGGTG 194 Query 411 CGCTCTAACCACCTGAGCTAATAGCCCTTACTCCGAACCGAACTCA--GTTTGAAA 464 Sbjct 193 CGCTCTAACCACCTGAGCTAATAGCCCATACTCCGAACC-AATTGATAGTTTGAAA 139 Score = 124 bits (67), Expect = 5e-25 Identities = 77/81 (95%), Gaps = 4/81 (4%) Strand=Plus/Minus Query 197 TTTTGGTGGAGGTAAGCGGACTCGAACCGCTGACATCCTGC-TTGCAAAG-CAGGCGCTC 254 Sbjct 335 TTTTGGTGGAGGTAAGCGGACTCGAACCGCTGACATC-TGCCTTGCAAAGGCAG-CGCTC 278 Query 255 TACCAACTGAGCTATACCCCC 275 Sbjct 277 TACCAACTGAGCTATACCCCC KK 16S-23S M. chthonoplastes SA18 EF M. chthonoplastes WW8 EF M. chthonoplastes GNL1 EF Microcoleus chthonoplastes WW12 EF M. chthonoplastes EDA EF Arthrospira platensis M2 AY Geitlerinema cf. acuminatum CCALA Phormidium autumnale UTCC 579 AM Bal pse Uncultured cyanobacterium isolate DGGE gel band 3 16S ribosomal RNA gene, partial sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; and 23S ribosomal RNA gene, partial sequence Length=588 Score = 259 bits (140), Expect = 1e-65 Identities = 153/159 (96%), Gaps = 2/159 (1%) Strand=Plus/Plus
127 105 Query 100 TGGGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGATAAGGGTGAGGTCCCTGGTTC 159 Sbjct 231 TGGGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGATAAGGGTGAGGTCCCTGGTTC 290 Query 160 AAGTCCAGGATGGCCCACTA--TGGGGGTATAGCTCAGTTGGTAGAGCGCCTGCTTTGCA 217 Sbjct 291 GAGTCCAGGATGGCCCACTGCGTGGGGGTATAGCTCAGTTGGTAGAGCGCCTGCTTTGCA 350 Query 218 CGCAGGAAGTCAGCGGTTCGAGTCCGCTTACCTCCACCA 256 Sbjct 351 AGCAGGATGTCAGCGGTTCGAGTCCGCTTACCTCCACCA 389 R Uncultured cyanobacterium isolate DGGE gel band 3 16S ribosomal RNA gene, partial sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; and 23S ribosomal RNA gene, partial sequence Length=588 Score = 248 bits (134), Expect = 2e-62 Identities = 151/159 (94%), Gaps = 2/159 (1%) Strand=Plus/Minus Query 193 TGGGGGAGGTAAGCGGACTCGAACCGCTGACTTCCTGCGTGCAAAGCAGGCGCTCTACCA 252 Sbjct 389 TGGTGGAGGTAAGCGGACTCGAACCGCTGACATCCTGCTTGCAAAGCAGGCGCTCTACCA 330 Query 253 ACTGAGCTATACCCCCA--TAGTGGGCCATCCTGGACTTGAACCAGGGACCTCACCCTTA 310 Sbjct 329 ACTGAGCTATACCCCCACGCAGTGGGCCATCCTGGACTCGAACCAGGGACCTCACCCTTA 270 Query 311 TCAGGGGTGCGCTCTAACCACCTGAGCTAATACCCCCCA 349 Sbjct 269 TCAGGGGTGCGCTCTAACCACCTGAGCTAATAGCCCCCA 231 Phormidium sp. OL 05 AM Phormidium sp. OL 32 AM Phormidium sp. OL M10 AM Phormidium sp. OL S5 AM Oscillatoriales cyanobacterium Gollwi Leptolyngbya sp. PCC 8936 AM Uncultured cyanobacterium DGGE gel ba... Cyanobacterium sp. LLi5 DQ Cyanobacterium sp. MBIC10216 AM Cyanobacterium aponinum ETS-03 AM Phormidium persicinum SAG AM Bal pse 16S-23S Plectonema terebrans CCAP 1463/4 AY76... Leptolyngbya sp. Kovacik 1986/5a EU52... Oscillatoria sp. CCMEE 416 AM ZD
128 106 Geitlerinema cf. acuminatum CCALA S-23S ribosomal RNA intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence Length=444 Score = 420 bits (227), Expect = 4e-114 Identities = 357/416 (85%), Gaps = 25/416 (6%) Strand=Plus/Plus Query 21 AAAAATAATTAGGGAAGAGAAAGGGGTCAGGACCTAGGTCGGTCGAGGATTAGAAGCAGG 80 Sbjct 32 AAAAGTAATTAGGTAAGAGAAA-GGGTCA-GACCTAGGTCGGTCGAGGATTAGAAGCAGG 89 Query 81 aaaaaaaaagggga-ttcgggtcggagg-atgggctattagctcttttggttagagcgca 138 Sbjct 90 -CTTTCAAACTCAACTTCGGTTCGGAGGTATGGGCTATTAGCTCAGGTGGTTAGAGCGCA 148 Query 139 CCCCTGATAAGGGGGAGGGCCCTGGTTCGAGTCCCGGATGGCCCACCTAAACCCaaaaaa 198 Sbjct 149 CCCCTGATAAGGGTGAGGTCCCTGGTTCGAGTCCAGGATGGCCCACCTAAACCCAAAAGA 208 Query 199 accctgcgtaagggaataaagtgggggtatagctcagttggtagagcgctgcctttgcaa 258 Sbjct 209 ACCTTGAAAAAGGG-A-AAA-TGGGGGTATAGCTCAGTTGGTAGAGCGCTGCCTTTGCAA 265 Query 259 GGCAGATGTCAGCGGTTCGAGTCCGCTTACCTCCAGTGGAGGAAAAACCAAGAGTCAGCA 318 Sbjct 266 GGCAGATGTCAGCGGTTCGAGTCCGCTTACCTCCWGTGGAGGAAAAAT-AAGAATCAGCA 324 Query A-C--TCAT-AAG-T-AA-TGAGACTGCTGGCTAA-CTTGCCAGCCCGAACCTTGA 366 Sbjct 325 CCTCATCAATCATCAAAATGAAATGAGACTGCTGG-TAGTCTTACCAGCCAGAACCTTGA 383 Query 367 AAAGTACCTAGCACTAAGTCT-AAAAAGTCTAGGGAAAATA-ATCTAACAGACATT 420 Sbjct 384 AAAGTGCATAGCAATAAGTCTTAAAAAGTCTAGGGAAAA-AGATCAAACAGACATT 438 R Geitlerinema cf. acuminatum CCALA S-23S ribosomal RNA intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence Length=444 Score = 327 bits (177), Expect = 3e-86 Identities = 363/448 (81%), Gaps = 34/448 (7%) Strand=Plus/Minus Query 24 AATGTCTGTTTGATCTTTTTCCCTAGAC-TTTT-AGACTTATGGCTATGTACTTTCCAAT 81 Sbjct 438 AATGTCTGTTTGATCTTTTTCCCTAGACTTTTTAAGACTTATTGCTATGCACTTTTCAAG 379 Query 82 GTaaaaaaaGGCAAG-TTAGCCAG-AATCTCA-TT-A--CT--T-A-TG--G-GTTGCTA 128 Sbjct 378 GTTCTGGCTGGTAAGACTA-CCAGCAGTCTCATTTCATTTTGATGATTGATGAGGTGCTG 320 Query 129 ACTCTTGGTTTTTCCTCCACTGGGGGTAAGCGGACTCGAACCGCTGACATCTGCCTTGCA 188 Sbjct 319 ATTCTT-ATTTTTCCTCCACWGGAGGTAAGCGGACTCGAACCGCTGACATCTGCCTTGCA 261 Query 189 AAGGAAGCGCTCTACCAACTGAGATATACCCCCACTTTATTCCCTTACGCA-GGtttttt 247 Sbjct 260 AAGGCAGCGCTCTACCAACTGAGCTATACCCCCA-TTT-T-CCCTTTTTCAAGGTTCTTT 204 Query 248 tgggttaaggtgggccatcctggactcgacccagacacctcaccctgataaggggtg-gc 306 Sbjct 203 TGGGTTTAGGTGGGCCATCCTGGACTCGAACCAGGGACCTCACCCTTATCAGGGGTGCGC 144 Query 307 CTCTAACCCCCTGAGCTAATAccccccaccccc-aacccccA-TGGAATTTGAAAGCCAG 364
129 107 Sbjct 143 -TCTAACCACCTGAGCTAATAGCCCATACCTCCGAACCGA-AGTTGAGTTTGAAAGCCTG 86 Query 365 CTTCTAATCCCCGACCGACCAAGGGCTGACCCTTTCTCTAACCAAAAttt-tttt-ttCG 422 Sbjct 85 CTTCTAATCCTCGACCGACCTAGGTCTGACCCTTTCTCTTACCTAA-TTACTTTTGTTCG 27 Query 423 GGCA--GAGGGGGTTT-GGTCCCCCTAA 447 Sbjct 26 GT-ATTGAGGGG-TTTAGGTCTCCCTAA Oscillatoria sp. PCC 8989 FJ Oscillatoriales cyanobacterium Schier Phormidiaceae cyanobacterium SAG Phormidium sp. OL S6 AM Phormidium willei BDU AJ Leptolyngbya valderiana BDU AJ9... Uncultured Planktothricoides sp. clon... ZD 16S-23S Geitlerinema cf. acuminatum CCALA Şif Geitlerinema sp. PCC S-23S ribosomal RNA intergenic spacer, complete sequence Length=624 Score = 1003 bits (543), Expect = 0.0 Identities = 555/560 (99%), Gaps = 3/560 (0%) Strand=Plus/Plus Query 15 AGGA-AT-AAT-AGCCCACTCAATTGAGGTCAACCTAGGTCGTGCGAAGAGTATTCGGTC 71 Sbjct 65 AGGATATAAATAAGCCCACTCAATTGAGGTCAACCTAGGTCGTGCGAAGAGTATTCGGTC 124 Query 72 GTGTCTTTCAAACGAACGAGGATTCGGAATCTCGGTTTTCAGTTTTTGAAAACTCGACGA 131 Sbjct 125 GTGTCTTTCAAACGAACGAGGATTCGGAATCTCGGTTTTCAGTTTTTGAAAACTCGACGA 184 Query 132 GGCCGAAAAACGTGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGATAAGGGTGAGG 191 Sbjct 185 GGCCGAAAAACGTGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGATAAGGGTGAGG 244 Query 192 TCCCTGGTTCAAGTCCAGGATGGCCCACCTGATTCAGCGAGCAGTGAGCAGCAAACAGTG 251 Sbjct 245 TCCCTGGTTCAAGTCCAGGATGGCCCACCTGATTCAGCGAGCAGTGAGCAGCAAACAGCG 304 Query 252 AGCAGTGAAAACTGATGACTGGTGACTGGTGACTGTTAACTGAAATCGTGGGGGTATAGC 311 Sbjct 305 AGCAGTGAAAACTGATGACTGGTGACTGGTGACTGTTAACTGAAATCGTGGGGGTATAGC 364 Query 312 TCAGTTGGTAGAGCGCCTGCTTTGCAAGCAGGATGTCAGCGGTTCGAGTCCGCTTACCTC 371 Sbjct 365 TCAGTTGGTAGAGCGCCTGCTTTGCAAGCAGGATGTCAGCGGTTCGAGTCCGCTTACCTC 424 Query 372 CACGCTCGTCTCGTTGCCACTCGAACCTCGAATCCCAAACGATTCTAATCGGAAAGGGAG 431 Sbjct 425 CACGCTCGTCTCGTCGCCACTCGAACCTCGAATCCCAAACGATTCTAATCGGAAAGGGAG 484 Query 432 TTCAGCACCCGAGAGAGCTCGGACTGCTGGACGTACCGTCCAGTAAGAACCTTGAAAACT 491 Sbjct 485 TTCAGCACCCGAGAGAGCTCGGACTGCTGGACGTACCGTCCAGTAAGAACCTTGAAAACT 544 Query 492 GCATAGTCATCTCTAAACAAAGTAAAGCTAGGGAAAGCCGAGACCTCGCGAATAGAGGTC 551
130 108 Sbjct 545 GCATAGTCATCTCTAAACAAAGTAAAGCTAGGGAAAGCCGAGACCTCGCGAATAGAGGTC 604 Query 552 AAGTTACAAAGGGCTAACGG 571 Sbjct 605 AAGTTACAAAGGGCTAACGG 624 R Geitlerinema sp. PCC S-23S ribosomal RNA intergenic spacer, complete sequence Length=624 Score = 1048 bits (567), Expect = 0.0 Identities = 590/601 (98%), Gaps = 2/601 (0%) Strand=Plus/Minus Query 3 CTTTTTAATTTTACCTCTATGCGCGAGGTCTCGGCTTTCCCTAGCTTTACTTTGTTTAGA 62 Sbjct 615 CTTTGTAACTTGACCTCTATTCGCGAGGTCTCGGCTTTCCCTAGCTTTACTTTGTTTAGA 556 Query 63 GATGACTATGCAGTTTTCAAGGTTCTTACTGGACGGTACGTCCAGCAGTCCGAGCTCTCT 122 Sbjct 555 GATGACTATGCAGTTTTCAAGGTTCTTACTGGACGGTACGTCCAGCAGTCCGAGCTCTCT 496 Query 123 CGGGTGCTGAACTCCCTTTCCGATTAGAATCGTTTGGGATTCGAGGTTCGAGTGGCAACG 182 Sbjct 495 CGGGTGCTGAACTCCCTTTCCGATTAGAATCGTTTGGGATTCGAGGTTCGAGTGGCGACG 436 Query 183 AGACGAGCGGGGAGGTAAGCGGACTCGAACCGCTGACATCCTGCTTGCAAAGCAGGCGCT 242 Sbjct 435 AGACGAGCGTGGAGGTAAGCGGACTCGAACCGCTGACATCCTGCTTGCAAAGCAGGCGCT 376 Query 243 CTACCAACTGAGCTATACCCCCACGATTTCAGTTAACAGTCACCAGTCACCAGTCATCAG 302 Sbjct 375 CTACCAACTGAGCTATACCCCCACGATTTCAGTTAACAGTCACCAGTCACCAGTCATCAG 316 Query 303 TTTTCACTGCTCACTGTTTGCTGCTCACTGCTCGCTGAATCAGGGGGGCCATCCTGGACT 362 Sbjct 315 TTTTCACTGCTCGCTGTTTGCTGCTCACTGCTCGCTGAATCAGGTGGGCCATCCTGGACT 256 Query 363 TGAACCAGGGACCTCACCCTTATCAGGGGTGCGCTCTAACCACCTGAGCTAATAGCCCAC 422 Sbjct 255 TGAACCAGGGACCTCACCCTTATCAGGGGTGCGCTCTAACCACCTGAGCTAATAGCCCAC 196 Query 423 GTTTTTCGGCCTCGTCGAGTTTTCAAAAACTGAAAACCGAGATTCCGAATCCTCGTTCGT 482 Sbjct 195 GTTTTTCGGCCTCGTCGAGTTTTCAAAAACTGAAAACCGAGATTCCGAATCCTCGTTCGT 136 Query 483 TTGAAAGACACGACCGAATACTCTTCGCACGACCTAGGTTGACCTCAATTGAGTGGGCTT 542 Sbjct 135 TTGAAAGACACGACCGAATACTCTTCGCACGACCTAGGTTGACCTCAATTGAGTGGGCTT 76 Query 543 ATTTATATCCTTCACTCAATCGGGGAACGGTCTCCCTAAAAGGAGGTGATCCCAGCC-CA 601 Sbjct 75 ATTTATATCCTTCACTCAATTGGGGAACGGTCTCCCTAAAAGGAGGTGATCC-AGCCACA 17 Query 602 C 602 Sbjct 16 C 16
131 Leptolyngbya valderiana BDU AJ Leptolyngbya valderiana BDU AJ... Geitlerinema sp. Flo1 FJ Geitlerinema sp. PCC 7105 FJ Sif 16S-23S Scytonema hyalinum FI-8A AF Nodularia harveyana CCAP 1452/1 AY Bal ip Leptolyngbya frigida ANT.LH S-23S ribosomal RNA intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence Length=530 Score = 340 bits (184), Expect = 5e-90 Identities = 221/237 (93%), Gaps = 10/237 (4%) Strand=Plus/Plus Query 41 AGAGTTGGTCATCCC-AAGGTCGTTCGGAT-TACGGATG-TGGCTTCCAAACTGTTTGGT 97 Sbjct 60 AGAGTTGGCCATCCCAAAGGTCGTTCGGATATA-TG-TGTTGGCTTCCAAACTATTTGGT 117 Query 98 TCGGTGT-ATGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGATAAGGGTGAGGTCC 156 Sbjct 118 TCGGT-TCATGGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGATAAGGGTGAGGTCC 176 Query 157 CTGGTTCGAGTCCAGGATGGCCCACCTTTACATATTGATAAGGGGGTTTAGCTCAGTTGG 216 Sbjct 177 CTGGTTCGAGTCCAGGATGGCCCACCTTTACAT-T--ACAAGGGGGTTTAGCTCAGTTGG 233 Query 217 TAGAGCGCCTGCTTTGCAAGCAGGATGTCAGCGGTTCGAGTCCGCTAACCTCCACTG 273 Sbjct 234 TAGAGCGCCTGCTTTGCAAGCAGGATGTCAACGGTTCGAGTCCGTTAACCTCCACTG 290 R Leptolyngbya frigida ANT.LH S-23S ribosomal RNA intergenic spacer, partial sequence; and trna-ile and trna-ala genes, complete sequence Length=530 Score = 340 bits (184), Expect = 5e-90 Identities = 221/237 (93%), Gaps = 10/237 (4%) Strand=Plus/Minus Query 341 CAGTGGAGGTTAGCGGACTCGAACCGCTGACATCCTGCTTGCAAAGCAGGCGCTCTACCA 400 Sbjct 290 CAGTGGAGGTTAACGGACTCGAACCGTTGACATCCTGCTTGCAAAGCAGGCGCTCTACCA 231 Query 401 ACTGAGCTAAACCCCCTTATCAATATGTAAAGGTGGGCCATCCTGGACTCGAACCAGGGA 460 Sbjct 230 ACTGAGCTAAACCCCCTTGT--A-ATGTAAAGGTGGGCCATCCTGGACTCGAACCAGGGA 174 Query 461 CCTCACCCTTATCAGGGGTGCGCTCTAACCACCTGAGCTAATAGCCCAT-ACACCGAACC 519 Sbjct 173 CCTCACCCTTATCAGGGGTGCGCTCTAACCACCTGAGCTAATAGCCCATGA-ACCGAACC 115 Query 520 AAACAGTTTGGAAGCCA-CATCCGTA-ATCCGAACGACCTT-GGGATGACCAACTCT 573 Sbjct 114 AAATAGTTTGGAAGCCAACA-CA-TATATCCGAACGACCTTTGGGATGGCCAACTCT 60
132 Leptolyngbya sp. Kovacik 1990/54 EU Leptolyngbya sp. Kovacik 1990/37 EU52... Leptolyngbya sp. LLi18 DQ Leptolyngbya sp. CCALA 094 AM Phormodium pristleyi TM4LUS131 EU Leptolyngbya foveolarum P-BJ1 EU L. foveolarum P-BJ1 EU Leptolyngbya frigida ANT.LH70.1 AY Leptolyngbya frigida ANT.MANNING.1 AY... Bal ip 16S-23S 0.05 KY Phormidium autumnale UTCC S rrna gene (partial), 23S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 Length=593 Sort alignments for this subject sequence by: E value Score Percent identity Query start position Subject start position Score = 172 bits (93), Expect = 2e-39 Identities = 109/116 (93%), Gaps = 4/116 (3%) Strand=Plus/Minus Query 356 TTTGGGGTTCAGGTGGGCCATCCTGGACTTGAACCAGGGACCTCACCCTTATCAGGGGTG 415 Sbjct 252 TTTGGGGTT-AGGTGGGCCATCCTGGACTTGAACCAGGGACCTCACCCTTATCAGGGGTG 194 Query 416 CGCTCTAACCACCTGAGCTAATAGCCCTTACTCCGAACCGAACTCA--GTTTGAAA 469 Sbjct 193 CGCTCTAACCACCTGAGCTAATAGCCCATACTCCGAACC-AATTGATAGTTTGAAA 139 Score = 124 bits (67), Expect = 5e-25 Identities = 77/81 (95%), Gaps = 4/81 (4%) Strand=Plus/Minus Query 202 TTTTGGTGGAGGTAAGCGGACTCGAACCGCTGACATCCTGC-TTGCAAAG-CAGGCGCTC 259 Sbjct 335 TTTTGGTGGAGGTAAGCGGACTCGAACCGCTGACATC-TGCCTTGCAAAGGCAG-CGCTC 278 Query 260 TACCAACTGAGCTATACCCCC 280 Sbjct 277 TACCAACTGAGCTATACCCCC 257 R Phormidium autumnale UTCC S rrna gene (partial), 23S rrna gene (partial), trna-ile gene, trna-ala gene and ITS1 Length=593 Sort alignments for this subject sequence by: E value Score Percent identity
133 111 Query start position Subject start position Score = 172 bits (93), Expect = 2e-39 Identities = 109/116 (93%), Gaps = 4/116 (3%) Strand=Plus/Minus Query 356 TTTGGGGTTCAGGTGGGCCATCCTGGACTTGAACCAGGGACCTCACCCTTATCAGGGGTG 415 Sbjct 252 TTTGGGGTT-AGGTGGGCCATCCTGGACTTGAACCAGGGACCTCACCCTTATCAGGGGTG 194 Query 416 CGCTCTAACCACCTGAGCTAATAGCCCTTACTCCGAACCGAACTCA--GTTTGAAA 469 Sbjct 193 CGCTCTAACCACCTGAGCTAATAGCCCATACTCCGAACC-AATTGATAGTTTGAAA 139 Score = 124 bits (67), Expect = 5e-25 Identities = 77/81 (95%), Gaps = 4/81 (4%) Strand=Plus/Minus Query 202 TTTTGGTGGAGGTAAGCGGACTCGAACCGCTGACATCCTGC-TTGCAAAG-CAGGCGCTC 259 Sbjct 335 TTTTGGTGGAGGTAAGCGGACTCGAACCGCTGACATC-TGCCTTGCAAAGGCAG-CGCTC 278 Query 260 TACCAACTGAGCTATACCCCC 280 Sbjct 277 TACCAACTGAGCTATACCCCC Microcoleus chthonoplastes WW12 EF Microcoleus chthonoplastes WW8 EF M. chthonoplastes SA18 EF M. chthonoplastes GNL1 EF Phormidium autumnale UTCC 579 AM Geitlerinema cf. acuminatum CCALA KY 16S-23S 100 Arthrospira platensis M2 AY U sal Uncultured marine bacterium clone SPOTSAPR01_14m146 16S ribosomal RNA gene, partial sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; and 23S ribosomal RNA gene, partial sequence Length=851 Score = 141 bits (76), Expect = 6e-30 Identities = 139/166 (83%), Gaps = 17/166 (10%) Strand=Plus/Plus Query 154 GTAGCTCAGGTGGTTAGAGCGCACGCCTGATAAGCGTGAGGTCGGAGGTTCAAGTCCTCC 213 Sbjct 256 GTAGCTCAGGTGGTTAGAGCGCACGCCTGATAAGCGTGAGGTCGGAGGTTCAAGTCCTCC 315 Query 214 -CAAGCCCACAATGG-TAACCG--AGG-GGC-TATAGCTCAGCTGGCTAGAGCAC-TGCC 266 Sbjct 316 TCGA-CCCACCATGGATTACCCTTAAAAGGCGT-TAGCTCAGCTGGG-AGAGCACCTGC- 371 Query 267 TTTGCAAGGCAGGGG-TCCGGGGTTCGAATCCCCG-TA-GCTCCAC 309 Sbjct 372 TTTGCAAG-CAGGGGGTCATCGGTTCGA-TCCC-GATACGCTCCAC 414
134 112 R Uncultured marine bacterium clone SPOTSAPR01_14m146 16S ribosomal RNA gene, partial sequence; 16S-23S ribosomal RNA intergenic spacer, complete sequence; and 23S ribosomal RNA gene, partial sequence Length=851 Score = 141 bits (76), Expect = 6e-30 Identities = 139/166 (83%), Gaps = 17/166 (10%) Strand=Plus/Minus Query 413 GTGGAGC-TA-CGGGGATTCGAACCCCGGA-CCCCTGCCTTGCAAAGGCA-GTGCTCTAG 468 Sbjct 414 GTGGAGCGTATC-GGGA-TCGAACCGATGACCCCCTG-CTTGCAAA-GCAGGTGCTCT-C 360 Query 469 CCAGCTGAGCTATA-GCC-CCT--CGGTTA-CCATTGTGGGCTTG-GGAGGACTTGAACC 522 Sbjct 359 CCAGCTGAGCTA-ACGCCTTTTAAGGGTAATCCATGGTGGG-TCGAGGAGGACTTGAACC 302 Query 523 TCCGACCTCACGCTTATCAGGCGTGCGCTCTAACCACCTGAGCTAC 568 Sbjct 301 TCCGACCTCACGCTTATCAGGCGTGCGCTCTAACCACCTGAGCTAC 256 U ip Phormidium persicinum SAG S rrna gene (partial), ITS1, 23S rrna gene (partial), trna-ile gene and trna-ala gene, strain SAG Length=625 Score = 246 bits (133), Expect = 9e-62 Identities = 149/156 (95%), Gaps = 4/156 (2%) Strand=Plus/Plus Query 122 GGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGATAAGGGTGAGGTCCCTGGTTCGAG 181 Sbjct 258 GGGCTATTAGCTCAGGTGGTTAGAGCGCACCCCTGATAAGGGTGAGGTCCCTGGTTCGAG 317 Query 182 TCCAGGATGGCCCACTGAGGGGGTATAGCTCAGTTGGTAGAGCGC-TGCCTTTGCAAGGC 240 Sbjct 318 TCCAGGATGGCCCACTGAGGGGGTATAGCTCAGTTGGTAGAGCGCCTGC-TTTGCAAG-C 375 Query 241 AGAA-GCCAGCGGTTCGAGCCCGCTTACCTCCACCA 275 Sbjct 376 AGGATGTCAGCGGTTCGAGTCCGCTTACCTCCACCA 411 R Phormidium persicinum SAG S rrna gene (partial), ITS1, 23S rrna gene (partial), trna-ile gene and trna-ala gene, strain SAG Length=625 Score = 246 bits (133), Expect = 9e-62 Identities = 149/156 (95%), Gaps = 4/156 (2%) Strand=Plus/Minus Query 194 TGGTGGAGGTAAGCGGGCTCGAACCGCTGGC-TTCTGCCTTGCAAAGGCA-GCGCTCTAC 251 Sbjct 411 TGGTGGAGGTAAGCGGACTCGAACCGCTGACATCCTG-CTTGCAAA-GCAGGCGCTCTAC 354 Query 252 CAACTGAGCTATACCCCCTCAGTGGGCCATCCTGGACTCGAACCAGGGACCTCACCCTTA 311 Sbjct 353 CAACTGAGCTATACCCCCTCAGTGGGCCATCCTGGACTCGAACCAGGGACCTCACCCTTA 294 Query 312 TCAGGGGTGCGCTCTAACCACCTGAGCTAATAGCCC 347 Sbjct 293 TCAGGGGTGCGCTCTAACCACCTGAGCTAATAGCCC 258
135 Phormidium persicinum SAG AM Leptolyngbya sp. PCC 8936 AM Uncultured cyanobacterium DGGE gel ba... Oscillatoriales cyanobacterium Gollwi... Phormidium sp. OL 05 AM Phormidium sp. OL 32 AM Uncultured Chroococcidiopsis sp. clon... Uncultured Chroococcidiopsis sp. clon(2) U ip 16S-23S 0.05 Çizelge 4.4: Pc βf-αr dizi analizi sonuçları Primerler Organizma Adı Maksimum 1 DF (Pc βf-αr) U pse Agmenellum quadruplicatum phycocyanin alpha and beta subunit genes, complete cds 1 DR (Pc βf-αr) U pse Agmenellum quadruplicatum phycocyanin alpha and beta subunit genes, complete cds 2DF (Pc βf-αr) U osc Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds 2 DR (Pc βf-αr) U osc Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds 3 DF (Pc βf-αr) KK Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin 3 DR (Pc βf-αr) KK Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin 4 DF (Pc βf-αr) Bal pse Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds 4 DR (Pc βf-αr) Bal pse - 5 DF (Pc βf-αr) ZD - 5 DR (Pc βf-αr) ZD - 6 DF (Pc βf-αr) Şif Geitlerinema sp. PCC 7105 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds 6 DR (Pc βf-αr) Şif Geitlerinema sp. PCC 7105 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds 7 DF (Pc βf-αr) Bal ip Uncultured cyanobacterium clone SY08PC-05 phycocyanin beta subunit (pcb) and phycocyanin alpha subunit (pca) genes, partial cds Tanımlanma %89 %90 %81 %82 %82 %80 %82 %99 % 99 %79 7 DR (Pc βf-αr) Bal ip Uncultured cyanobacterium clone SY08PC-05 phycocyanin beta subunit (pcb) and phycocyanin alpha subunit (pca) genes, partial cds 8 DF (Pc βf-αr) KY Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds 8 DR (Pc βf-αr) KY Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds 9 DF (Pc βf-αr) U sal Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds 9 DR (Pc βf-αr)u sal Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds 10 DF (Pc βf-αr)u ip Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds 10 DR (Pc βf-αr)u ip Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds %76 % 83 % 83 % 82 % 81 % 82 % 82 U pse Agmenellum quadruplicatum phycocyanin alpha and beta subunit genes, complete cds Length=1695
136 114 Score = 462 bits (250), Expect = 1e-126 Identities = 333/373 (89%), Gaps = 5/373 (1%) Strand=Plus/Plus Query 336 GTAAGACAATCTTTTTAAAGAAAATTATTGATAATCCATTAGGGAGATAGTTTGACAATG 395 Sbjct 951 GTAAGACGCTC-TTTTAAAGAAAATTATTGATAATCCA-TAGGGAGATAATCTGACAATG 1008 Query 396 AAAACCCCTCTTACCGAAGCAGTAGCGA-TCGCCGACTCTCAAGGTCGTTTCCTCAGCAA 454 Sbjct 1009 AAAACCCCTCTTACCGAAGCAGTAGC-ACTCGCTGATTCTCAAGGCCGTTTCCTCAGCAA 1067 Query 455 CACTGAGCTCCAGTACCTCTATGGTCGTCTCCGTCAAGGTGCTTTTGCTCTTGATGCAGC 514 Sbjct 1068 CACTGAGCTCCAGTACCTCTATGGTCGTCTTCGTCAAGGTGCTTTCGCCCTTGAAGCGGC 1127 Query 515 TCAAGCTTTGACTGCAAAGGCTGACACTCTCGTTAGCGGTGCAGCTCAAGCTGTTTACAG 574 Sbjct 1128 TCAAACGTTGACTGCAAAAGCTGACACCCTCGTTAATGGTGCTGCTCAAGCGGTTTACAG 1187 Query 575 CAAGTTCCCTTACACAACTAGCACTCCCGGTAACAACTTTGCTGCTGACCAACGCGGTAA 634 Sbjct 1188 CAAGTTCCCCTACACCACCAGCACTCCTGGCAACAACTTCGCTGCTGACCAGCGCGGTAA 1247 Query 635 GGACAAGTGTGCTCGTGACATCGGTTACTACCTCCGCATGATCACTTATTGTTTAGTTGC 694 Sbjct 1248 AGACAAGTGTGCTCGTGACATCGGTTACTACCTCCGCATGGTTACCTACTGCCTAGTTGC 1307 Query 695 TGGGGGGTACTGG 707 Sbjct 1308 TGGTGG-TACTGG 1319 R Agmenellum quadruplicatum phycocyanin alpha and beta subunit genes, complete cds Length=1695 Score = 416 bits (225), Expect = 8e-113 Identities = 291/323 (90%), Gaps = 4/323 (1%) Strand=Plus/Minus Query 17 CGATGTCACGAGCACACTTGTCCTTACCGCGTTGGTCAGCAGCAAAGTTGTTACCGGGAG 76 Sbjct 1270 CGATGTCACGAGCACACTTGTCTTTACCGCGCTGGTCAGCAGCGAAGTTGTTGCCAGGAG 1211 Query 77 TGCTAGTTGTGTAAGGGAACTTGCTGTAAACAGCTTGAGCTGCACCGCTAACGAGAGTGT 136 Sbjct 1210 TGCTGGTGGTGTAGGGGAACTTGCTGTAAACCGCTTGAGCAGCACCATTAACGAGGGTGT 1151 Query 137 CAGCCTTTGCAGTCAAAGCTTGAGCTGCATCAAGAGCAAAAGCACCTTGACGGAGACGAC 196 Sbjct 1150 CAGCTTTTGCAGTCAACGTTTGAGCCGCTTCAAGGGCGAAAGCACCTTGACGAAGACGAC 1091 Query 197 CATAGAGGTACTGGAGCTCAGTGTTGCTGAGGAAACGACCTTGAGAGTCGGCGA-TCGCT 255 Sbjct 1090 CATAGAGGTACTGGAGCTCAGTGTTGCTGAGGAAACGGCCTTGAGAATCAGCGAGT-GCT 1032 Query 256 ACTGCTTCGGTAAGAGGGGTTTTCATTGTCAAACTATCTCCCTAATGGATTATCAATAAT 315 Sbjct 1031 ACTGCTTCGGTAAGAGGGGTTTTCATTGTCAGATTATCTCCCTA-TGGATTATCAATAAT 973 Query 316 TTTCTTTAAAAAGATTGTCTTAC 338 Sbjct 972 TTTCTTTAAAA-GAGCGTCTTAC 951 U osc Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750
137 115 Score = 259 bits (140), Expect = 2e-65 Identities = 275/338 (81%), Gaps = 17/338 (5%) Strand=Plus/Plus Query 404 TTAGGAGATTTCC-AGAACAATGAAAACCCCCATTACTGAAGCTATTGCTGCAGCTGACG 462 Sbjct 355 TTAGGAGA-ATCCGAGAACAATGAAGACCCCTATCACCGAAGCAATTTCTTCTGCTGATT 413 Query 463 CTCAAGGACG-TTTCGTCAGCAACACCGAACTGCAATTCGAG-A-ATGGTCGTTTAGACC 519 Sbjct 414 CCCAAGGACGCTTTC-TGAGCAACAGCGAACTGCAA-TC-TGTACGTGGCCGCTTCGAGC 470 Query 520 GCGCTGCTGCCAGCATGGAAGCGGCTCGCGCTTTAAC-CGGCAAATCTAGCGAACTGA-T 577 Sbjct 471 GCGCTACTGCCAGCATGGAAGCGGCTCGCGCTTTAACTCAG-AATTCTAGCTC-CTTAGT 528 Query 578 CAAT-GGCGCTGCTAACGCTGATTATCAGAAGTTCCCTTACACCACCCAAATGCAAGGGG 636 Sbjct 529 -AAGCGGTGCGGCTAACGCCGTTTACCAAAAATTCCCTTACACCACCCAAATGCAAGGGC 587 Query 637 CTAACTACGCTTACGACAAACGCGG-AAAGATAAATGTTCTCGTGACATCGGCCACTATC 695 Sbjct 588 CTAACTACGCTTACGACCAACGCGGTAAAGACAAATGCGCTCGTGACATCGGTCACTACC 647 Query 696 TCCGCATGGTGACATACTGCTT-AGTTGCTGGGGGTAC 732 Sbjct 648 TGCGCATGGTGACTTACTGCCTGATTT-CTGGTGGTAC 684 R Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750 Score = 244 bits (132), Expect = 4e-61 Identities = 244/296 (82%), Gaps = 15/296 (5%) Strand=Plus/Minus Query 14 GTGCCCGATGTCACGAG-ACATTTATCTTTACCGCGTTTGTCGTAAGCGTAGTTAGCCCC 72 Sbjct 643 GTGACCGATGTCACGAGCGCATTTGTCTTTACCGCGTTGGTCGTAAGCGTAGTTAGGCCC 584 Query 73 TTGCATTTGGGTGGTGTAAGGGAACTTCTGATAAACAGCGTTAGCAGCGCC-ATTGA-TC 130 Sbjct 583 TTGCATTTGGGTGGTGTAAGGGAATTTTTGGTAAACGGCGTTAGCCGCACCGCTT-ACTA 525 Query 131 AGTTCGCTAGATTTACCG-GTTAAAGCGCGAGCCGCTTCCATGCTGGCAGCAGCGCGGTC 189 Sbjct 524 AG-GAGCTAGAATT-CTGAGTTAAAGCGCGAGCCGCTTCCATGCTGGCAGTAGCGCGCTC 467 Query 190 TAAACGACCAT-T-CACGAATTGCAGTTCGGTGTTGCTGACGAAA-CGTCCTTGAGCGTC 246 Sbjct 466 GAAGCGGCCACGTACA-GA-TTGCAGTTCGCTGTTGCTCA-GAAAGCGTCCTTGGGAATC 410 Query 247 AGCTGCAGCAATAGCTTCAGTAATGGGGGTTTTCATTGTTCT-GGAAATCTCCTAA 301 Sbjct 409 AGCAGAAGAAATTGCTTCGGTGATAGGGGTCTTCATTGTTCTCGGAT-TCTCCTAA 355 KK Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=578 Score = 198 bits (107), Expect = 3e-47 Identities = 180/215 (83%), Gaps = 5/215 (2%) Strand=Plus/Plus Query 300 AACGATGAAAACTCCGATTACTGAAGCAATCGCCGCAGCTGATACCCAAGGTCGTTTCCT 359
138 116 Sbjct 327 AAC-ATGAAGACCCCAATTACCGAAGCGATCGCAGCAGCTGATACCCAAGGCCGTTTTCT 385 Query 360 CG-GCAACACCGAACTCCAAGCGGTTAACGGTCGTTTCGAGCGTGCTGTTGCCAGCATGG 418 Sbjct 386 -GAGCAACACCGAACTGCAAGCTGTTCGCGGTCGGTTAGAACGCGCTGGCGCTAGCATGG 444 Query 419 AAGCTGCTCGCGCTCTGACCAACAAAGCTCAAA-GCTTGATCGATGGTGCTGCTAACGCG 477 Sbjct 445 AAGCTGCTCGCGCTCTGACCAACAACGCGCAAAAGCTAGTT-GACGGTGCAGCTAACGCG 503 Query 478 GTTTATCAGAAGTTCCCCTACACCACTCAAATGCA 512 Sbjct 504 GTGTATCAAAAGTACCCCTACACCACCCAACTGCA 538 R Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=578 Score = 198 bits (107), Expect = 3e-47 Identities = 180/215 (83%), Gaps = 5/215 (2%) Strand=Plus/Minus Query 73 TGCATTTGAGTGGTGTAGGGGAACTTCTGATAAACCGCGTTAGCAGCACCATCGATCAAG 132 Sbjct 538 TGCAGTTGGGTGGTGTAGGGGTACTTTTGATACACCGCGTTAGCTGCACCGTC-AACTAG 480 Query 133 C-TTTGAGCTTTGTTGGTCAGAGCGCGAGCAGCTTCCATGCTGGCAACAGCACGCTCGAA 191 Sbjct 479 CTTTTGCGCGTTGTTGGTCAGAGCGCGAGCAGCTTCCATGCTAGCGCCAGCGCGTTCTAA 420 Query 192 ACGACCGTTAACCGCTTGGAGTTCGGTGTTGC-CGAGGAAACGACCTTGGGTATCAGCTG 250 Sbjct 419 CCGACCGCGAACAGCTTGCAGTTCGGTGTTGCTC-AGAAAACGGCCTTGGGTATCAGCTG 361 Query 251 CGGCGATTGCTTCAGTAATCGGAGTTTTCATCGTT 285 Sbjct 360 CTGCGATCGCTTCGGTAATTGGGGTCTTCAT-GTT 327 Bal pse Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=578 Score = 171 bits (92), Expect = 6e-39 Identities = 170/207 (82%), Gaps = 8/207 (3%) Strand=Plus/Plus Query 308 AACGATGAAAACTCCGATTACTGAAGCAATCGCCGCAGCTGATACCCAAGGTCGTTTCCT 367 Sbjct 327 AAC-ATGAAGACCCCAATTACCGAAGCGATCGCAGCAGCTGATACCCAAGGCCGTTTTCT 385 Query 368 CG-GCAACACCGAACTCCAAGCGGTTAACGGTCGTTTCGAGCGTGCTGTTGCCAGCATGG 426 Sbjct 386 -GAGCAACACCGAACTGCAAGCTGTTCGCGGTCGGTTAGAACGCGCTGGCGCTAGCATGG 444 Query 427 AAGCTGCTCGCGCTCTGACCAACAAAAGCTCAAA-GCTTGATCGAAGGTGCTGCTAAACC 485 Sbjct 445 AAGCTGCTCGCGCTCTGACCAACAAC-GCGCAAAAGCTAGTT-GACGGTGCAGCTAA-CG 501 Query 486 CGGTTTATCAGAAATTTCCCCTACACC 512 Sbjct 502 CGGTGTATCA-AAAGTACCCCTACACC 527 Şif Geitlerinema sp. PCC 7105 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=731
139 117 Score = 1085 bits (587), Expect = 0.0 Identities = 596/600 (99%), Gaps = 1/600 (0%) Strand=Plus/Plus Query 20 GTTTTCTCTGGCGATGCCAGCGTTCTCGACGATCGTTGCTTGAACGGTCTGCGCGAAACC 79 Sbjct 126 GTTTTCTCTGGCGATGCCAGCGTTCTCGACGATCGTTGCTTGAACGGTCTGCGCGAAACC 185 Query 80 TACGTCGCTCTCGGAACTCCCGGTGCGTCCGTAGCCGTCGGCGTTCAAAAAATGAAAGAA 139 Sbjct 186 TACGTCGCTCTCGGAACTCCCGGTGCGTCCGTAGCCGTCGGCGTTCAAAAAATGAAAGAA 245 Query 140 GCTGCCCTGCAAATCGTGGGCGATCCCGCTGGTATCACCCCCGGAGATTGCAGCAGCCTG 199 Sbjct 246 GCTGCCCTGCAAATCGTGGGCGATCCCGCTGGTATCACCCCTGGAGATTGCAGCAGCCTG 305 Query 200 ATGTCTGAAATCGGAACCTACTTCGATCGCGCGGCTGCCGCTGTTGCGTAAATCGCACAG 259 Sbjct 306 ATGTCTGAAATCGGAACCTACTTCGATCGCGCGGCTGCCGCTGTTGCGTAAATCGCACAG 365 Query 260 TAGCTCAGTTTCCTCGAGTCCGCTTAGCGGACATCTCAATTTGAAAATCGTTTTGACAGT 319 Sbjct 366 TAGCTCAGTTTCCTCGAGTCCGCTTAGCGGACATCTCAATTTGAAAATCGTTTTGACAGT 425 Query 320 CAAAAAGGAGATTACTCAACAATGAAGACCCCGTTAACCGAAGCAGTCGCTGCAGCAGAT 379 Sbjct 426 CAAAAAGGAGATTACTCAACAATGAAGACCCCGTTAACCGAAGCAGTCGCTGCAGCAGAT 485 Query 380 TCTCAAGGTCGCTTCCTCAGCAGCACCGAACTCCAAACCGCGTTCGGTCGTTTCCGTCAA 439 Sbjct 486 TCTCAAGGTCGCTTCCTCAGCAGCACCGAACTCCAAACCGCGTTCGGTCGTTTCCGTCAA 545 Query 440 GCCAATGCCGGCTTGCAAGCTGCCCGTCAACTGACCGACAAATCTCAGTCTCTGATCGAC 499 Sbjct 546 GCCAATGCCGGCTTGCAAGCTGCCCGTCAACTGACCGACAAATCTCAGTCTCTGATCGAC 605 Query 500 GGTGCCGCTCAAGCGGTGTATAACAAATTCCCTTACACCACTCAAATGCAAGGGGCGAAC 559 Sbjct 606 GGTGCCGCTCAAGCGGTGTATAACAAATTCCCTTACACCACTCAAATGCAAGGGGCAAAC 665 Query 560 TACGCTTCTAGCCCCGAAGGCAAAGCGAAGTGCTCTCGCGACATCGGCTACTACCTGCGT 619 Sbjct 666 TACGCTTCTAGCCCTGAAGGCAAAGCGAAGTGCTCTCGCGACATCGGCTACTACC-GCGT 724 R Geitlerinema sp. PCC 7105 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=731 Score = 1151 bits (623), Expect = 0.0 Identities = 629/632 (99%), Gaps = 0/632 (0%) Strand=Plus/Minus Query 14 GTAGCCGATGTCGCGAGAGCACTTCGCTTTGCCTTCGGGGCTAGAAGCGTAGTTCGCCCC 73 Sbjct 716 GTAGCCGATGTCGCGAGAGCACTTCGCTTTGCCTTCAGGGCTAGAAGCGTAGTTTGCCCC 657 Query 74 TTGCATTTGAGTGGTGTAAGGGAATTTGTTATACACCGCTTGAGCGGCACCGTCGATCAG 133 Sbjct 656 TTGCATTTGAGTGGTGTAAGGGAATTTGTTATACACCGCTTGAGCGGCACCGTCGATCAG 597 Query 134 AGACTGAGATTTGTCGGTCAGTTGACGGGCAGCTTGCAAGCCGGCATTGGCTTGACGGAA 193 Sbjct 596 AGACTGAGATTTGTCGGTCAGTTGACGGGCAGCTTGCAAGCCGGCATTGGCTTGACGGAA 537 Query 194 ACGACCGAACGCGGTTTGGAGTTCGGTGCTGCTGAGGAAGCGACCTTGAGAATCTGCTGC 253 Sbjct 536 ACGACCGAACGCGGTTTGGAGTTCGGTGCTGCTGAGGAAGCGACCTTGAGAATCTGCTGC 477 Query 254 AGCGACTGCTTCGGTTAACGGGGTCTTCATTGTTGAGTAATCTCCTTTTTGACTGTCAAA 313 Sbjct 476 AGCGACTGCTTCGGTTAACGGGGTCTTCATTGTTGAGTAATCTCCTTTTTGACTGTCAAA 417
140 118 Query 314 ACGATTTTCAAATTGAGATGTCCGCTAAGCGGACTCGAGGAAACTGAGCTACTGTGCGAT 373 Sbjct 416 ACGATTTTCAAATTGAGATGTCCGCTAAGCGGACTCGAGGAAACTGAGCTACTGTGCGAT 357 Query 374 TTACGCAACAGCGGCAGCCGCGCGATCGAAGTAGGTTCCGATTTCAGACATCAGGCTGCT 433 Sbjct 356 TTACGCAACAGCGGCAGCCGCGCGATCGAAGTAGGTTCCGATTTCAGACATCAGGCTGCT 297 Query 434 GCAATCTCCGGGGGTGATACCAGCGGGATCGCCCACGATTTGCAGGGCAGCTTCTTTCAT 493 Sbjct 296 GCAATCTCCAGGGGTGATACCAGCGGGATCGCCCACGATTTGCAGGGCAGCTTCTTTCAT 237 Query 494 TTTTTGAACGCCGACGGCTACGGACGCACCGGGAGTTCCGAGAGCGACGTAGGTTTCGCG 553 Sbjct 236 TTTTTGAACGCCGACGGCTACGGACGCACCGGGAGTTCCGAGAGCGACGTAGGTTTCGCG 177 Query 554 CAGACCGTTCAAGCAACGATCGTCGAGAACGCTGGCATCGCCAGAGAAAACTGCGTAGGT 613 Sbjct 176 CAGACCGTTCAAGCAACGATCGTCGAGAACGCTGGCATCGCCAGAGAAAACTGCGTAGGT 117 Query 614 GACGTAGCGCAGGATGATTTCCATGTCGCGTA 645 Sbjct 116 GACGTAGCGCAGGATGATTTCCATGTCGCGTA 85 Bal İp Uncultured cyanobacterium clone SY08PC-05 phycocyanin beta subunit (pcb) and phycocyanin alpha subunit (pca) genes, partial cds Length=660 Score = 150 bits (81), Expect = 9e-33 Identities = 233/303 (76%), Gaps = 24/303 (7%) Strand=Plus/Plus Query 393 ATGAAGACCCCGATTACTGAAGCGATCGCCGCAGCAGATACCCAAGGCCG-TTTTCTCAG 451 Sbjct 367 ATGAAAACACCAATTACCGAAGCAATCGCATCTGCTGATACCCAAGGACGCTTTT-TAAG 425 Query 452 CAACACCGAACTGCAAGCTGTTAACGGTCGCT-TTGAGCGTGCCGCTGCCAGCATGGAAG 510 Sbjct 426 CAACACCGAATTACAAGCAGTTAACGGTCG-TCTAGTTCGTGCCGCTGCCAGCATGGAAG 484 Query 511 CTGCTCGTGCGCTGACCAAC-AATGCCCAA--GCTCTGATCGACGGTGCTGCAAACGCCG 567 Sbjct 485 CTGCTCGTG-GTTTAACAGCTAACGCTCAAAAG-T-TGATTGACGGTGCTACCAGCGCTG 541 Query 568 TGTATCAG-AAGTTCCCCTACACCACC-C-AGATGCAGGGTTCAAACTATGC-C-TCCGA 622 Sbjct 542 TATA-CAGCAAATTCCCTTACACCACCTCTACA--CAAGG--CAATCAATTCGCATTTGA 596 Query 623 TGCTCGCGGCAA-GTCCAAGTGCGCTCGTGACATCGG-CTACTACCTGCGGATCATCACC 680 Sbjct 597 TCCTCGTGGTAAAG-CTAAGTGCGCTCGTGACGTTGGTC-ACTACCTCCGCATCATCACC 654 Query 681 TAC 683 Sbjct 655 TAC 657 R Uncultured cyanobacterium clone SY08PC-05 phycocyanin beta subunit (pcb) and phycocyanin alpha subunit (pca) genes, partial cds Length=660 Score = 134 bits (72), Expect = 9e-28 Identities = 164/207 (79%), Gaps = 12/207 (5%) Strand=Plus/Minus
141 119 Query 84 GGTGGTGTAGGGGAACTT-CTGATACACGGCGTTTGCAGCACCGTCGATCAGAGC--TTG 140 Sbjct 567 GGTGGTGTAAGGGAATTTGCTG-TATACAGCGCTGGTAGCACCGTCAATCA-A-CTTTTG 511 Query 141 GGCATT-GTTGGTCAGCGCACGAGCAGCTTCCATGCTGGCAGCGGCACGCTCAA-AGCGA 198 Sbjct 510 AGCGTTAGCTGTTAAAC-CACGAGCAGCTTCCATGCTGGCAGCGGCACGAACTAGA-CGA 453 Query 199 CCGTTAACAGCTTGCAGTTCGGTGTTGCTGAGAAAA-CGGCCTTGGGTATCTGCTGCGGC 257 Sbjct 452 CCGTTAACTGCTTGTAATTCGGTGTTGCTTA-AAAAGCGTCCTTGGGTATCAGCAGATGC 394 Query 258 GATCGCTTCAGTAATCGGGGTCTTCAT 284 Sbjct 393 GATTGCTTCGGTAATTGGTGTTTTCAT 367 KY Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=578 Score = 198 bits (107), Expect = 3e-47 Identities = 180/215 (83%), Gaps = 5/215 (2%) Strand=Plus/Plus Query 307 AACGATGAAAACTCCGATTACTGAAGCAATCGCCGCAGCTGATACCCAAGGTCGTTTCCT 366 Sbjct 327 AAC-ATGAAGACCCCAATTACCGAAGCGATCGCAGCAGCTGATACCCAAGGCCGTTTTCT 385 Query 367 CG-GCAACACCGAACTCCAAGCGGTTAACGGTCGTTTCGAGCGTGCTGTTGCCAGCATGG 425 Sbjct 386 -GAGCAACACCGAACTGCAAGCTGTTCGCGGTCGGTTAGAACGCGCTGGCGCTAGCATGG 444 Query 426 AAGCTGCTCGCGCTCTGACCAACAAAGCTCAAA-GCTTGATCGATGGTGCTGCTAACGCG 484 Sbjct 445 AAGCTGCTCGCGCTCTGACCAACAACGCGCAAAAGCTAGTT-GACGGTGCAGCTAACGCG 503 Query 485 GTTTATCAGAAGTTCCCCTACACCACTCAAATGCA 519 Sbjct 504 GTGTATCAAAAGTACCCCTACACCACCCAACTGCA 538 R Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=578 Score = 198 bits (107), Expect = 3e-47 Identities = 180/215 (83%), Gaps = 5/215 (2%) Strand=Plus/Minus Query 77 TGCATTTGAGTGGTGTAGGGGAACTTCTGATAAACCGCGTTAGCAGCACCATCGATCAAG 136 Sbjct 538 TGCAGTTGGGTGGTGTAGGGGTACTTTTGATACACCGCGTTAGCTGCACCGTC-AACTAG 480 Query 137 C-TTTGAGCTTTGTTGGTCAGAGCGCGAGCAGCTTCCATGCTGGCAACAGCACGCTCGAA 195 Sbjct 479 CTTTTGCGCGTTGTTGGTCAGAGCGCGAGCAGCTTCCATGCTAGCGCCAGCGCGTTCTAA 420 Query 196 ACGACCGTTAACCGCTTGGAGTTCGGTGTTGC-CGAGGAAACGACCTTGGGTATCAGCTG 254 Sbjct 419 CCGACCGCGAACAGCTTGCAGTTCGGTGTTGCTC-AGAAAACGGCCTTGGGTATCAGCTG 361 Query 255 CGGCGATTGCTTCAGTAATCGGAGTTTTCATCGTT 289 Sbjct 360 CTGCGATCGCTTCGGTAATTGGGGTCTTCAT-GTT 327
142 120 Usal Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750 Score = 274 bits (148), Expect = 5e-70 Identities = 266/321 (82%), Gaps = 16/321 (4%) Strand=Plus/Plus Query 395 TTAGGAGATTTCC-AGAACAATGAAAACCCCCATTACTGAAGCTATTGCTGCAGCTGACG 453 Sbjct 355 TTAGGAGA-ATCCGAGAACAATGAAGACCCCTATCACCGAAGCAATTTCTTCTGCTGATT 413 Query 454 CTCAAGGACG-TTTCGTCAGCAACACCGAACTGCAATTCGTG-A-ATGGTCGTTTAGACC 510 Sbjct 414 CCCAAGGACGCTTTC-TGAGCAACAGCGAACTGCAA-TC-TGTACGTGGCCGCTTCGAGC 470 Query 511 GCGCTGCTGCCAGCATGGAAGCGGCTCGCGCTTTAAC-CGGTAAATCTAGCGATC-TGA- 567 Sbjct 471 GCGCTACTGCCAGCATGGAAGCGGCTCGCGCTTTAACTCAG-AATTCTAGC--TCCTTAG 527 Query 568 TCAAT-GGCGCTGCTAACGCTGTTTATCAGAAGTTCCCTTACACCACCCAAATGCAAGGG 626 Sbjct 528 T-AAGCGGTGCGGCTAACGCCGTTTACCAAAAATTCCCTTACACCACCCAAATGCAAGGG 586 Query 627 GCTAACTACGCTTACGACAAACGCGGTAAAGATAAATGTTCTCGTGACATCGGCCACTAT 686 Sbjct 587 CCTAACTACGCTTACGACCAACGCGGTAAAGACAAATGCGCTCGTGACATCGGTCACTAC 646 Query 687 CTCCGCATGGTGACATACTGC 707 Sbjct 647 CTGCGCATGGTGACTTACTGC 667 R Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750 Score = 219 bits (118), Expect = 3e-53 Identities = 237/292 (81%), Gaps = 17/292 (5%) Strand=Plus/Minus Query 16 CGATGTCACGAG-ACATTTATCTTTACCGCGTTTGCTCGTAAGCGTAGTTAGCCCCTTGC 74 Sbjct 638 CGATGTCACGAGCGCATTTGTCTTTACCGCG-TTGGTCGTAAGCGTAGTTAGGCCCTTGC 580 Query 75 ATTTGGGCGG-GGAAGGGAACTTCTGATAAACAGCGTTAGCAGCGCC-ATTGA-TCAGTT 131 Sbjct 579 ATTTGGGTGGTGTAAGGGAATTTTTGGTAAACGGCGTTAGCCGCACCGCTT-ACTAAG-G 522 Query 132 CGCTAGATTTACCG-GTTAAAGCGCGAGCCGCTTCCATGCTGGCAGCAGCGCGGTCTAAA 190 Sbjct 521 AGCTAGAATT-CTGAGTTAAAGCGCGAGCCGCTTCCATGCTGGCAGTAGCGCGCTCGAAG 463 Query 191 CGACCAT-T-CACGAATTGCAGTTCGGTGTTGCTGACGAAA-CGTCCTTGAGCGTCAGCT 247 Sbjct 462 CGGCCACGTACA-GA-TTGCAGTTCGCTGTTGCTCA-GAAAGCGTCCTTGGGAATCAGCA 406 Query 248 GCAGCAATAGCTTCAGTAATGGGGGTTTTCATTGTTCT-GGAAATCTCCTAA 298 Sbjct 405 GAAGAAATTGCTTCGGTGATAGGGGTCTTCATTGTTCTCGGAT-TCTCCTAA 355 U ip Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750
143 121 Score = 279 bits (151), Expect = 1e-71 Identities = 280/340 (82%), Gaps = 18/340 (5%) Strand=Plus/Plus Query 400 TTAGGAGATTTCC-AGAACAATGAAAACCCCCATTACTGAAGCTATTGCTGCAGCTGACG 458 Sbjct 355 TTAGGAGA-ATCCGAGAACAATGAAGACCCCTATCACCGAAGCAATTTCTTCTGCTGATT 413 Query 459 CTCAAGGACG-TTTCGTCAGCAACACCGAACTGCAATTCGTG-A-ATGGTCGTTTAGACC 515 Sbjct 414 CCCAAGGACGCTTTC-TGAGCAACAGCGAACTGCAA-TC-TGTACGTGGCCGCTTCGAGC 470 Query 516 GCGCTGCTGCCAGCATGGAAGCGGCTCGCGCTTTAAC-CGGTAAATCTAGCGAACTGA-T 573 Sbjct 471 GCGCTACTGCCAGCATGGAAGCGGCTCGCGCTTTAACTCAG-AATTCTAGCTC-CTTAGT 528 Query 574 CAAT-GGCGCTGCTAACGCTGTTTATCAGAAGTTCCCTTACACCACCCAAATGCAAGGGG 632 Sbjct 529 -AAGCGGTGCGGCTAACGCCGTTTACCAAAAATTCCCTTACACCACCCAAATGCAAGGGC 587 Query 633 CTAACTACGCTTACGACAAACGCGGTAAAGATAAATGTTCTCGTGACATCGGCCACTATC 692 Sbjct 588 CTAACTACGCTTACGACCAACGCGGTAAAGACAAATGCGCTCGTGACATCGGTCACTACC 647 Query 693 TCCGCATGGTGACATACTGCTT-AGTTTGCTGGTGGGTAC 731 Sbjct 648 TGCGCATGGTGACTTACTGCCTGA-TTT-CTGGTGG-TAC 684 R Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750 Score = 237 bits (128), Expect = 7e-59 Identities = 239/291 (82%), Gaps = 14/291 (4%) Strand=Plus/Minus Query 21 CGATGTCACGAGAACATTTATCTTTACCGCGTTTGTCGTAAGCGTAGTTAGCCCCTTGCA 80 Sbjct 638 CGATGTCACGAGCGCATTTGTCTTTACCGCGTTGGTCGTAAGCGTAGTTAGGCCCTTGCA 579 Query 81 TTTGGGCGGTGTAAGGGAACTTCTGATAAACAGCGTTAGCAGCGCC-ATTGA-TCAGTTC 138 Sbjct 578 TTTGGGTGGTGTAAGGGAATTTTTGGTAAACGGCGTTAGCCGCACCGCTT-ACTAAG-GA 521 Query 139 GCTAGATTTACCG-GTTAAAGCGCGAGCCGCTTCCATGCTGGCAGCAGCGCGGTCTAAAC 197 Sbjct 520 GCTAGAATT-CTGAGTTAAAGCGCGAGCCGCTTCCATGCTGGCAGTAGCGCGCTCGAAGC 462 Query 198 GACCAT-T-CACGAATTGCAGTTCGGTGTTGCTGACGAAA-CGTCCTTGAGCGTCAGCTG 254 Sbjct 461 GGCCACGTACA-GA-TTGCAGTTCGCTGTTGCTCA-GAAAGCGTCCTTGGGAATCAGCAG 405 Query 255 CAGCAATAGCTTCAGTAATGGGGGTTTTCATTGTTCT-GGAAATCTCCTAA 304 Sbjct 404 AAGAAATTGCTTCGGTGATAGGGGTCTTCATTGTTCTCGGAT-TCTCCTAA 355
144 122 Çizelge 4.5: F-R dizi analizi sonuçları Primerler Organizma Adı Maksimum Tanımlanma 1 EF (F-R) U pse - 1 ER (F-R) U pse Planktothrix sp. PCC 7811 phycocyanin B subunit and phycocyanin A %79 subunit genes, partial cds 2 EF (F-R) U osc - 2 ER (F-R)U osc Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; %80 and phycocyanin alpha subunit gene, partial cds 3 EF (F-R) KK Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin %84 alpha subunit (cpca) genes, partial cds 3 ER (F-R)KK Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin % 82 alpha subunit (cpca) genes, partial cds 4 EF (F-R) Bal pse - 4 ER (F-R)Bal pse - 5 EF (F-R) ZD Arthrospira platensis FACHB341 cpc operon, partial sequence % 90 5 ER (F-R)ZD Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; %79 and phycocyanin alpha subunit gene, partial cds 6 EF (F-R) Şif Geitlerinema sp. PCC 7105 phycocyanin beta subunit (cpcb) and %98 phycocyanin alpha subunit (cpca) genes, partial cds 6 ER (F-R)Şif Geitlerinema sp. PCC 7105 phycocyanin beta subunit (cpcb) and % 99 phycocyanin alpha subunit (cpca) genes, partial cds 7 EF (F-R) Bal ip Uncultured cyanobacterium clone SY08PC-05 phycocyanin beta subunit % 78 (pcb) and phycocyanin alpha subunit (pca) genes, partial cds 7 ER (F-R)Bal ip Uncultured cyanobacterium clone SY08PC-05 phycocyanin beta subunit %84 (pcb) and phycocyanin alpha subunit (pca) genes, partial cds 8 EF (F-R) KY Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin % 84 alpha subunit (cpca) genes, partial cds 8 ER (F-R)KY Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin % 82 alpha subunit (cpca) genes, partial cds 9 EF (F-R) U sal Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; % 80 and phycocyanin alpha subunit gene, partial cds 9 ER (F-R)U sal Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; % 81 and phycocyanin alpha subunit gene, partial cds 10 EF (F-R) U ip - 10 ER (F-R) U ip - U Pse CTCGAGTGGCGGGGCTCAAGTCGGTCGTCGCTGCGATGGTGCCCCCAA ACATGGAAAGAAGCTGTTTCAAGTGGGGGGGCCCCCGGTATCCTCCTG GCGATTGCTCTGCGCCGATGCCATAAATCACCAGCTACTTTGATCGTG CTGCTTCTGCTGTTGGCTAAAGCCATTCTGGCCTACCAATTGAAGCTCA AAAATCTCACAAACACCTTGTAGGAAAACTATTTCAGGAAATATACAA ACAATGAAAACCCCGATTACCGAAGCGATCGCGGCAGCCAATACCCA AGGCCGTTTTCTTGGCAACCCCGAATTACAAGCAGTTGATGGTCGTTA CACGCGTGCAGCCCCTAGCATGGAAGCTGCTCGCAAACTGACTGACAA CGCTCAAAGCTTGATTAACAGCGCAGCTCAAGCTGTGTACTCCAAGTT CCCCTACACCCCCCAAATGCTTTTTTCTTTAAA R Planktothrix sp. PCC 7811 phycocyanin B subunit and phycocyanin A subunit genes, partial cds Length=662 Score = 135 bits (73), Expect = 2e-28 Identities = 171/216 (79%), Gaps = 16/216 (7%) Strand=Plus/Minus Query 254 TTAGCCAACAGCAGAAGCAGC-ACGATCAAAGTAGCTGGGGAGTTCAGACA-TCAGCGCA 311 Sbjct 264 TTAGGCGACAGCAGCCGCAGCGA-GATCAAAGTAGCCGGCGATTTCAGA-AGCCAGGGCA 207
145 123 Query 312 -GAGCAATCGCCAGGAGTGATACC-GGGAGGATCGTTAGCAACTTTG--ATAGCAGCTTC 367 Sbjct 206 CTA-CAATCTCCAGGAGTAATTCCTGCG-GGATCGTTAACAA--TAGCCAAAGCTGCATC 151 Query 368 TTTC-TTCTTGCTGACACCATTGGCAACAGAAGCACCAGGAACACCCAGAGCTTGGTAGG 426 Sbjct 150 TTTCATT-TTGCCAACGCCAACGGCGACGGAAGCACCAGGAACGCCCAGAGCCAGGTAGG 92 Query 427 TTTCACGCAGACCGTTCAAGCAACGATC-TTA-GAA 460 Sbjct 91 TTTCACGCAGACCGTTCAAGCAACGATCATTAAGAA 56 U Osc Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750 Score = 119 bits (64), Expect = 2e-23 Identities = 137/171 (80%), Gaps = 10/171 (5%) Strand=Plus/Minus Query 43 GCTAGATTTACCG-GTTAAAGCGCGAGCCGCTTCCATGCTGGAAGCAGCGCGGGCTAAAC 101 Sbjct 520 GCTAGAATT-CTGAGTTAAAGCGCGAGCCGCTTCCATGCTGGCAGTAGCGCGCTCGAAGC 462 Query 102 GACCAT-T-CACGAATTGCAGTTCGGTGTTGCTGACGAAA-CGTCCTTGAGCGTCAGCTG 158 Sbjct 461 GGCCACGTACA-GA-TTGCAGTTCGCTGTTGCTCA-GAAAGCGTCCTTGGGAATCAGCAG 405 Query 159 CAGCAATAGCTTCAGTAATGGGGGTTTTCATTGTTCT-GGAAATCTCCTAA 208 Sbjct 404 AAGAAATTGCTTCGGTGATAGGGGTCTTCATTGTTCTCGGAT-TCTCCTAA 355 KK Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=578 Score = 204 bits (110), Expect = 4e-49 Identities = 181/215 (84%), Gaps = 5/215 (2%) Strand=Plus/Plus Query 233 AACGATGAAAACTCCGATTACTGAAGCAATCGCCGCAGCTGATACCCAAGGTCGTTTCCT 292 Sbjct 327 AAC-ATGAAGACCCCAATTACCGAAGCGATCGCAGCAGCTGATACCCAAGGCCGTTTTCT 385 Query 293 CG-GCAACACCGAACTCCAAGCGGTTAACGGTCGTTTCGAGCGTGCTGTTGCCAGCATGG 351 Sbjct 386 -GAGCAACACCGAACTGCAAGCTGTTCGCGGTCGGTTAGAACGCGCTGGCGCTAGCATGG 444 Query 352 AAGCTGCTCGCGCTCTGACCAACAAAGCTCAAA-GCTTGATCGATGGTGCTGCTAACGCG 410 Sbjct 445 AAGCTGCTCGCGCTCTGACCAACAACGCGCAAAAGCTAGTT-GACGGTGCAGCTAACGCG 503 Query 411 GTTTATCAGAAGTTCCCCTACACCACCCAAATGCA 445 Sbjct 504 GTGTATCAAAAGTACCCCTACACCACCCAACTGCA 538 R Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=578 Score = 156 bits (84), Expect = 1e-34 Identities = 149/180 (82%), Gaps = 6/180 (3%) Strand=Plus/Minus
146 124 Query 20 CGCGATAGCAGCACCATCGATC-AGC-TTTGAGCTTTGTTGGTCAGAGCGCGAGCAGCTT 77 Sbjct 503 CGCGTTAGCTGCACCGTC-AACTAGCTTTTGCGCGTTGTTGGTCAGAGCGCGAGCAGCTT 445 Query 78 CCATGCTGGCAACAGCACGCTCGAAACGACCGTTAACCGCTTGGAGTTCGGTGTTGC-CG 136 Sbjct 444 CCATGCTAGCGCCAGCGCGTTCTAACCGACCGCGAACAGCTTGCAGTTCGGTGTTGCTC- 386 Query 137 AGGAAACGACCTTGGGTATCAGCTGCGGCGATTGCTTCAGTAATCGGAGTTTTCATCGTT 196 Sbjct 385 AGAAAACGGCCTTGGGTATCAGCTGCTGCGATCGCTTCGGTAATTGGGGTCTTCAT-GTT 327 ZD Arthrospira platensis FACHB341 cpc operon, partial sequence Length=2086 Score = 67.6 bits (36), Expect = 6e-08 Identities = 47/52 (90%), Gaps = 2/52 (3%) Strand=Plus/Plus Query 426 CAAGCTGTGTACTCCAAGTTCCCCTACACCACCCAAATGCAGGGGACCCTAA 477 Sbjct 1226 CAAGCAGTGTACAACAAGTTCCCCTACACCACCCAAATGCAGGG-ACC-TAA 1275 RSpirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750 Score = 143 bits (77), Expect = 1e-30 Identities = 167/209 (79%), Gaps = 12/209 (5%) Strand=Plus/Minus Query 264 TTTAGCCAACAGCAGAAGCAGCACGATCAAAGTAGCTGGCGAGTTCAGACA-TCA--GCG 320 Sbjct 255 TTTAGGCAACAGCAGCAGCAGCGCGATCGAAGTAGCTAGCAACTTCAGACATTAATTG-G 197 Query 321 CAAAGCAATCGCC-AGGAGTGATACCGGCAGGATCGTTAGCAACTTTGATAGCAGCTTCT 379 Sbjct 196 C--TGCAATCCCCTTGG-GTGATGCCGTTGGGGTCGTTAGCAATTTTAACCGCAGCGTCT 140 Query 380 TTCATCTT-GCTGACACCATTGGCAACAGAAGCACCAGGAACACCCAGAGCTTGGTAGGT 438 Sbjct 139 TTCATTTTGGC-AACGCCAGCCGCAACGGAAGCACCAGGAACACCCAGAGCTTGGTAGGT 81 Query 439 TTCACGCAGACCGTTCAAGCAACGAATCT 467 Sbjct 80 TTCACGCAGAC-GTTTAAACAACGA-TCT 54 Şif Geitlerinema sp. PCC 7105 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=731 Score = 806 bits (436), Expect = 0.0 Identities = 453/460 (98%), Gaps = 5/460 (1%) Strand=Plus/Plus Query 28 ACT-CCGG-GCGTCCGTAGCCGTCGGCGTTC-AAAAATGAAAGAAGCTGCCCTGC-AATC 83 Sbjct 201 ACTCCCGGTGCGTCCGTAGCCGTCGGCGTTCAAAAAATGAAAGAAGCTGCCCTGCAAATC 260 Query 84 GTGGGCGATCCCGCTGGTATCACCCCCGGAGATTGCAGCAGCCTGATGTCTGAAATCGGA 143 Sbjct 261 GTGGGCGATCCCGCTGGTATCACCCCTGGAGATTGCAGCAGCCTGATGTCTGAAATCGGA 320 Query 144 ACCTACTTCGATCGCGCGGCTGCCGCTGTTGCGTAAATCGCACAGTAGCTCAGTTTCCTC 203
147 125 Sbjct 321 ACCTACTTCGATCGCGCGGCTGCCGCTGTTGCGTAAATCGCACAGTAGCTCAGTTTCCTC 380 Query 204 GAGTCCGCTTAGCGGACATCTCAATTTGAAAATCGTTTTGACAGTCAAAAAGGAGATTAC 263 Sbjct 381 GAGTCCGCTTAGCGGACATCTCAATTTGAAAATCGTTTTGACAGTCAAAAAGGAGATTAC 440 Query 264 TCAACAATGAAGACCCCGTTAACCGAAGCAGTCGCTGCAGCAGATTCTCAAGGTCGCTTC 323 Sbjct 441 TCAACAATGAAGACCCCGTTAACCGAAGCAGTCGCTGCAGCAGATTCTCAAGGTCGCTTC 500 Query 324 CTCAGCAGCACCGAACTCCAAACCGCGTTCGGTCGTTTCCGTCAAGCCAATGCCGGCTTG 383 Sbjct 501 CTCAGCAGCACCGAACTCCAAACCGCGTTCGGTCGTTTCCGTCAAGCCAATGCCGGCTTG 560 Query 384 CAAGCTGCCCGTCAACTGACCGACAAATCTCAGTCTCTGATCGACGGTGCCGCTCAAGCG 443 Sbjct 561 CAAGCTGCCCGTCAACTGACCGACAAATCTCAGTCTCTGATCGACGGTGCCGCTCAAGCG 620 Query 444 GTGTATAACAAATTCCCTTACACCACCCAAATGCA-GGGG 482 Sbjct 621 GTGTATAACAAATTCCCTTACACCACTCAAATGCAAGGGG 660 R Geitlerinema sp. PCC 7105 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=731 Score = 837 bits (453), Expect = 0.0 Identities = 463/467 (99%), Gaps = 3/467 (0%) Strand=Plus/Minus Query 7 CACCGTC-TGAGCGGCA-CGTCGATCAGAGACTGAGATTTGTCGGTCAGTTGACGGGCAG 64 Sbjct 623 CACCG-CTTGAGCGGCACCGTCGATCAGAGACTGAGATTTGTCGGTCAGTTGACGGGCAG 565 Query 65 CTTGCAAGCCGGCATTGGCTTGACGGAAACGACCGAACGCGGTTTGGAGTTCGGTGCTGC 124 Sbjct 564 CTTGCAAGCCGGCATTGGCTTGACGGAAACGACCGAACGCGGTTTGGAGTTCGGTGCTGC 505 Query 125 TGAGGAAGCGACCTTGAGAATCTGCTGCAGCGACTGCTTCGGTTAACGGGGTCTTCATTG 184 Sbjct 504 TGAGGAAGCGACCTTGAGAATCTGCTGCAGCGACTGCTTCGGTTAACGGGGTCTTCATTG 445 Query 185 TTGAGTAATCTCCTTTTTGACTGTCAAAACGATTTTCAAATTGAGATGTCCGCTAAGCGG 244 Sbjct 444 TTGAGTAATCTCCTTTTTGACTGTCAAAACGATTTTCAAATTGAGATGTCCGCTAAGCGG 385 Query 245 ACTCGAGGAAACTGAGCTACTGTGCGATTTACGCAACAGCGGCAGCCGCGCGATCGAAGT 304 Sbjct 384 ACTCGAGGAAACTGAGCTACTGTGCGATTTACGCAACAGCGGCAGCCGCGCGATCGAAGT 325 Query 305 AGGTTCCGATTTCAGACATCAGGCTGCTGCAATCTCCGGGGGTGATACCAGCGGGATCGC 364 Sbjct 324 AGGTTCCGATTTCAGACATCAGGCTGCTGCAATCTCCAGGGGTGATACCAGCGGGATCGC 265 Query 365 CCACGATTTGCAGGGCAGCTTCTTTCATTTTTTGAACGCCGACGGCTACGGACGCACCGG 424 Sbjct 264 CCACGATTTGCAGGGCAGCTTCTTTCATTTTTTGAACGCCGACGGCTACGGACGCACCGG 205 Query 425 GAGTTCCGAGAGCGACGTAGGTTTCGCGCAGACCGTTCAAGCAACGA 471 Sbjct 204 GAGTTCCGAGAGCGACGTAGGTTTCGCGCAGACCGTTCAAGCAACGA 158 Bal ip Uncultured cyanobacterium clone SY08PC-05 phycocyanin beta subunit (pcb) and phycocyanin alpha subunit (pca) genes, partial cds Length=660
148 126 Score = 128 bits (69), Expect = 3e-26 Identities = 163/207 (78%), Gaps = 12/207 (5%) Strand=Plus/Plus Query 325 ATGAAGACCCCGATTACTGAAGCGATCGCCGCAGCAAATACCCAAGGCCG-TTTTCTCAG 383 Sbjct 367 ATGAAAACACCAATTACCGAAGCAATCGCATCTGCTGATACCCAAGGACGCTTTT-TAAG 425 Query 384 CAACACCGAACTGCAAGCTGTTAACGGTCGCT-TTGAGCGTGCCGCTGCCAGCATGGAAG 442 Sbjct 426 CAACACCGAATTACAAGCAGTTAACGGTCG-TCTAGTTCGTGCCGCTGCCAGCATGGAAG 484 Query 443 CTGCTCGTGCGCTGACCAAC-AATGCCCAA--GCTCTGATCGACGGTGCTGCAAACGCCG 499 Sbjct 485 CTGCTCGTG-GTTTAACAGCTAACGCTCAAAAG-T-TGATTGACGGTGCTACCAGCGCTG 541 Query 500 TGTATCAG-AAGTTCCCCTACACCACC 525 Sbjct 542 TATA-CAGCAAATTCCCTTACACCACC 567 R Uncultured cyanobacterium clone SY08PC-05 phycocyanin beta subunit (pcb) and phycocyanin alpha subunit (pca) genes, partial cds Length=660 Score = 124 bits (67), Expect = 4e-25 Identities = 108/128 (84%), Gaps = 2/128 (1%) Strand=Plus/Minus Query 59 CACGAGCAGCTTCCATGCTGGCAGCGGCACGCTCAA-AGCGACCGTTAACAGCTTGCAGT 117 Sbjct 493 CACGAGCAGCTTCCATGCTGGCAGCGGCACGAACTAGA-CGACCGTTAACTGCTTGTAAT 435 Query 118 TCGGTGTTGCTGAAAAAACGGCCTTGGGTATCTGCTGCGGCGATCGCTTCAGTAATCGGG 177 Sbjct 434 TCGGTGTTGCTTAAAAAGCGTCCTTGGGTATCAGCAGATGCGATTGCTTCGGTAATTGGT 375 Query 178 GTCTTCAT 185 Sbjct 374 GTTTTCAT 367 KY Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=578 Score = 204 bits (110), Expect = 4e-49 Identities = 181/215 (84%), Gaps = 5/215 (2%) Strand=Plus/Plus Query 231 AACGATGAAAACTCCGATTACTGAAGCAATCGCCGCAGCTGATACCCAAGGTCGTTTCCT 290 Sbjct 327 AAC-ATGAAGACCCCAATTACCGAAGCGATCGCAGCAGCTGATACCCAAGGCCGTTTTCT 385 Query 291 CG-GCAACACCGAACTCCAAGCGGTTAACGGTCGTTTCGAGCGTGCTGTTGCCAGCATGG 349 Sbjct 386 -GAGCAACACCGAACTGCAAGCTGTTCGCGGTCGGTTAGAACGCGCTGGCGCTAGCATGG 444 Query 350 AAGCTGCTCGCGCTCTGACCAACAAAGCTCAAA-GCTTGATCGATGGTGCTGCTAACGCG 408 Sbjct 445 AAGCTGCTCGCGCTCTGACCAACAACGCGCAAAAGCTAGTT-GACGGTGCAGCTAACGCG 503 Query 409 GTTTATCAGAAGTTCCCCTACACCACCCAAATGCA 443 Sbjct 504 GTGTATCAAAAGTACCCCTACACCACCCAACTGCA 538
149 127 R Lyngbya nigra SC11 phycocyanin beta subunit (cpcb) and phycocyanin alpha subunit (cpca) genes, partial cds Length=578 Score = 154 bits (83), Expect = 4e-34 Identities = 149/180 (82%), Gaps = 7/180 (3%) Strand=Plus/Minus Query 19 CGCG-TAGCAGCACCATCGATC-AGC-TTTGAGCTTTGTTGGTCAGAGCGCGAGCAGCTT 75 Sbjct 503 CGCGTTAGCTGCACCGTC-AACTAGCTTTTGCGCGTTGTTGGTCAGAGCGCGAGCAGCTT 445 Query 76 CCATGCTGGCAACAGCACGCTCGAAACGACCGTTAACCGCTTGGAGTTCGGTGTTGC-CG 134 Sbjct 444 CCATGCTAGCGCCAGCGCGTTCTAACCGACCGCGAACAGCTTGCAGTTCGGTGTTGCTC- 386 Query 135 AGGAAACGACCTTGGGTATCAGCTGCGGCGATTGCTTCAGTAATCGGAGTTTTCATCGTT 194 Sbjct 385 AGAAAACGGCCTTGGGTATCAGCTGCTGCGATCGCTTCGGTAATTGGGGTCTTCAT-GTT 327 U sal Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750 Score = 172 bits (93), Expect = 2e-39 Identities = 197/245 (80%), Gaps = 15/245 (6%) Strand=Plus/Plus Query 327 TTAGGAGATTTCC-AGAACAATGAAAACCCCCATTACTGAAGCTATTGCTGCAGCTGACG 385 Sbjct 355 TTAGGAGA-ATCCGAGAACAATGAAGACCCCTATCACCGAAGCAATTTCTTCTGCTGATT 413 Query 386 CTCAAGGACG-TTTCGTCAGCAACACCGAACTGCAATTCGTG-A-ATGGTCGTTTAGACC 442 Sbjct 414 CCCAAGGACGCTTTC-TGAGCAACAGCGAACTGCAA-TC-TGTACGTGGCCGCTTCGAGC 470 Query 443 GCGCTGCTGCCAGCATGGAAGCGGCTCGCGCTTTAAC-CGGTAAATCTAGCGAACTGA-T 500 Sbjct 471 GCGCTACTGCCAGCATGGAAGCGGCTCGCGCTTTAACTCAG-AATTCTAGCTC-CTTAGT 528 Query 501 CAAT-GGCGCTGCTAACGCTGTTTATCAGAAGTTCCCTTACACCACCCAAATGCAGGGGA 559 Sbjct 529 -AAGCGGTGCGGCTAACGCCGTTTACCAAAAATTCCCTTACACCACCCAAATGCAAGGG- 586 Query 560 CCTAA 564 Sbjct 587 CCTAA 591 R Spirulina platensis phycocyanin beta subunit-like gene, partial sequence; and phycocyanin alpha subunit gene, partial cds Length=750 Score = 130 bits (70), Expect = 9e-27 Identities = 139/171 (81%), Gaps = 10/171 (5%) Strand=Plus/Minus Query 39 GCTAGATTTACCG-GTTAAAGCGCGAGCCGCTTCCATGCTGGCAGCAGCGCGGTCTAAAC 97 Sbjct 520 GCTAGAATT-CTGAGTTAAAGCGCGAGCCGCTTCCATGCTGGCAGTAGCGCGCTCGAAGC 462 Query 98 GACCAT-T-CACGAATTGCAGTTCGGTGTTGCTGACGAAA-CGTCCTTGAGCGTCAGCTG 154 Sbjct 461 GGCCACGTACA-GA-TTGCAGTTCGCTGTTGCTCA-GAAAGCGTCCTTGGGAATCAGCAG 405
150 128 Query 155 CAGCAATAGCTTCAGTAATGGGGGTTTTCATTGTTCT-GGAAATCTCCTAA 204 Sbjct 404 AAGAAATTGCTTCGGTGATAGGGGTCTTCATTGTTCTCGGAT-TCTCCTAA S rrna ve 16S-23S ITS primerleri dizi analizi sonuçunda birbirini doğrulamıştır. Fikosiyanin primerleri ile yapılan sonuçların bazıları dizi analizi için yeterli olmamıştır. Nübel et al., (1997) 16S rrna dizilerine dayanarak yayınladığı, siyanobakteriyel rrna gen parçalarının seçici alımı için doğal ve yapay kurulumda bir PCR yöntemi geliştirmiştir. Bu prosedürü DGGE ile birleştirerek, DNA moleküllerinin dizi-bağlı ayırımları tekniğini oluşturmuşlardır. Çevresel örnekler içindeki 16S rrna siyanobakteriyel genlerinin çeşitliliğini bu yöntem kullanılarak göstermiş ve izole edilen türlerin benzer olmadıkları kanıtlanmıştır. DGGE, PCR ürünlerinin DNA moleküllerinin tek bir homojen populasyonu içeriğinde tek bantlarından veya doğrudan dizisi olabilir, yaklaşık 700 nükleotidi olan 16S rrna genlerinin verimliliği hakkında da bilgi verdiği tanımlanmıştır (Nübel et al., 1997). Türlerin DGGE kullanılarak farklı türler olduğu gösterilmiştir (Şekil 4.19, 4.20, 4.21, 4.22, 4.23). Aynı primerler ile farklı bölgelerin çoğaltıldığı ve tek bir bazdaki değişimin farklılığı PCR-DGGE ile belirlenmiştir. Şekil te saflaştrılmayan karışık kültürlerden elde edilen sonuçlar gösterilmektedir. Belirli bir kommunite DNA sındaki 16S rrna tanımlı genler DGGE yöntemi ile farklılıkları gösterilmiştir. Hongmei ve arkadaşları 36 türün sekanslarının DGGE bantlarından farklı türler olduğunu gösterilmiştir. Filogenetik analizler, Calothrix, Cyanothece, Fischerella, Phormidium, Pleurocapsa, Oscillatoria ve Synechococcus cinsleri içinden, mezofilik karşılıklarından ayrılarak oluşan yeni termofilik nesilleri gösterilmiştir (Hongmei et al., 2005). Kısmen halofilik, tuza dayanıklı ve termofilik yeni bir siyanobakteri cinsine ait 4 tür üzerine 16S rrna gen sekansına dayanan filogenetik analizler bu türlerin yakından ilişkili (%99) olduklarını göstermiştir ve bu türler için Halomicronema excentrium önerilmiştir (Abed et al., 2002).
. M a b c d e f g h ı i j k l m n o Şekil 4.")
151 129 CYA 359F ve CYA 781R (a ve b) primerleri ve 40 nükleotidli Guanin Sitozin (GC) ce zengin primer dizisi ile çoğaltılan PCR ürünleri, DGGE de denaturasyon jeli % gradientte olacak şekilde hazırlanarak 80 V da 15 saat yürütülmüş ve görüntüleme cihazı yardımıyla bantlar görüntülenmiştir (Şekil 4.19, 4.20, 4.21, 4.22, 4.23, 4.24). M a b c d e f g h ı i j k l m n o Şekil 4. 19: DGGE de Şif, U pse, KK, KY türlerinin karşılaştırılması. Marker 100bp DNA ladder (1,517bp, 1,200bp, 1,000bp, 900bp, 800bp, 700bp, 600bp, bp, 400bp, 300bp, 200bp, 100bp), a; Şif 1 (CYA 359-CYA 781-a), b; Şif 2 (CYA 359-CYA 781-b), c; Şif 3 (CYA 359-CYA 781-a*), d; Şif 4 (CYA 359-CYA 781-b*); e; U pse 1 (CYA 359-CYA 781-a), f; U pse 2 (CYA 359-CYA 781-b), g; U pse 3 (CYA 359-CYA 781-a*), h; U pse 4 (CYA 359-CYA 781-b*); ı; KK 1 (CYA 359-CYA 781-a), i; KK 2 (CYA 359-CYA 781-b), j; KK 3 (CYA 359-CYA 781-a*), k; KK 4 (CYA 359-CYA 781-b*); l; KY 1 (CYA 359-CYA 781-a), m; KY 2 (CYA 359-CYA 781- b), n; KY 3 (CYA 359-CYA 781-a*), o; KY 4 (CYA 359-CYA 781-b*).
, KK 2 (CYA 359-CYA 781-b), KK 3 (CYA 359-CYA 781-a*), KK 4 (CYA 359-CYA 781-b*); ZD 1 (CYA 359-CYA 781-a), ZD 2 (CYA 359-CYA 781-b), ZD 3 (CYA 359-CYA 781-a*), ZD 4")
152 130 M Şekil 4.20: DGGE de KK, ZD, Şif türlerinin karşılaştırılması. Marker, KK 1 (CYA 359-CYA 781- a), KK 2 (CYA 359-CYA 781-b), KK 3 (CYA 359-CYA 781-a*), KK 4 (CYA 359-CYA 781-b*); ZD 1 (CYA 359-CYA 781-a), ZD 2 (CYA 359-CYA 781-b), ZD 3 (CYA 359-CYA 781-a*), ZD 4 (CYA 359-CYA 781-b*); Şif 1 (CYA 359-CYA 781-a), Şif 2 (CYA 359-CYA 781-b), Şif 3 (CYA 359-CYA 781-a*), Şif 4 (CYA 359-CYA 781-b*)
, b, KK 2 (CYA 359-CYA 781-b), c, KK 3 (CYA 359-CYA 781-a*), d, KK 4 (CYA 359-CYA 781- b*); e, U ip 1 (CYA 359-CYA 781-a), f, U ip 2 (CYA 359-CYA 781-b), g, U ip 3 (CYA")
153 131 a b c d e f g h ı i k l M Şekil 4.21: DGGE de KK, U ip, ZD türlerinin karşılaştırılması. a, KK 1 (CYA 359-CYA 781-a), b, KK 2 (CYA 359-CYA 781-b), c, KK 3 (CYA 359-CYA 781-a*), d, KK 4 (CYA 359-CYA 781- b*); e, U ip 1 (CYA 359-CYA 781-a), f, U ip 2 (CYA 359-CYA 781-b), g, U ip 3 (CYA 359-CYA 781-a*), h, U ip 4 (CYA 359-CYA 781-b*); ı, ZD 1 (CYA 359-CYA 781-a), i, ZD 2 (CYA 359- CYA 781-b), k, ZD 3 (CYA 359-CYA 781-a*), l, ZD 4 (CYA 359-CYA 781-b*), Marker(M).
, b, KY 2 (CYA 359-CYA 781-b), c, KY 3 (CYA 359-CYA 781-a*), d, KY 4 (CYA 359-CYA 781-b*); e, KK 1 (CYA 359-CYA 781-a), f, KK 2")
, l, Bal pse 4 (CYA 359- CYA 781-b*); m, Bal ip 1 (CYA 359-CYA 781-a), n, Bal ip 2 (CYA 359-CYA 781-b), o, Bal ip 3 (CYA 359-CYA")
154 132 a b c d e f g h ı i k l m n o p M Şekil 4.22: DGGE de KY, KK, Bal pse, Bal ip türlerinin karşılaştırılması. a, KY 1 (CYA 359- CYA 781-a), b, KY 2 (CYA 359-CYA 781-b), c, KY 3 (CYA 359-CYA 781-a*), d, KY 4 (CYA 359-CYA 781-b*); e, KK 1 (CYA 359-CYA 781-a), f, KK 2 (CYA 359-CYA 781-b), g, KK 3 (CYA 359-CYA 781-a*), h, KK 4 (CYA 359-CYA 781-b*); ı, Bal pse 1 (CYA 359-CYA 781-a), i, Bal pse 2 (CYA 359-CYA 781-b), k, Bal pse 3 (CYA 359-CYA 781-a*), l, Bal pse 4 (CYA 359- CYA 781-b*); m, Bal ip 1 (CYA 359-CYA 781-a), n, Bal ip 2 (CYA 359-CYA 781-b), o, Bal ip 3 (CYA 359-CYA 781-a*), p, Bal ip 4 (CYA 359-CYA 781-b*), Marker(M).
, b, Upse 2 (CYA 359-CYA 781-b), c, Upse 3 (CYA 359-CYA 781-a*), d, Upse 4 (CYA 359-CYA 781-b*); e, U osc 1 (CYA 359-CYA 781-a),")
, j, U ip 3 (CYA 359-CYA 781-a*), k, U ip 4 (CYA 359-CYA 781-b*); l, U sal 1 (CYA 359-CYA 781-a), m, U sal 2 (CYA 359-CYA 781-b), n, U sal 3 (CYA 359-CYA")
155 133 a b c d e f g h ı i j k l m n o M Şekil 4.23: DGGE de U pse, U osc, U ip, U sal türlerinin karşılaştırılması. a, Upse 1 (CYA 359- CYA 781-a), b, Upse 2 (CYA 359-CYA 781-b), c, Upse 3 (CYA 359-CYA 781-a*), d, Upse 4 (CYA 359-CYA 781-b*); e, U osc 1 (CYA 359-CYA 781-a), f, U osc 2 (CYA 359-CYA 781-b), g, U osc 3 (CYA 359-CYA 781-a*), h, U osc 4 (CYA 359-CYA 781-b*); ı, U ip 1 (CYA 359- CYA 781-a), i, U ip 2 (CYA 359-CYA 781-b), j, U ip 3 (CYA 359-CYA 781-a*), k, U ip 4 (CYA 359-CYA 781-b*); l, U sal 1 (CYA 359-CYA 781-a), m, U sal 2 (CYA 359-CYA 781-b), n, U sal 3 (CYA 359-CYA 781-a*), o, U sal 4 (CYA 359-CYA 781-b*), Marker (M)
, a, U (CYA 359-CYA 781-a), b, Ş (CYA 359-CYA 781-a), c, Z (CYA 359-CYA 781-a), d, K (CYA 359-CYA 781-a); e, U (CYA 359-CYA 781-b), f, Ş (CYA 359-CYA 781-b), g, Z (CYA")
, n, Z (CYA 359- CYA 781-b*), o, K (CYA 359-CYA 781-b*), p, U (CYA 359*-CYA 781-a*), r, Ş (CYA 359*- CYA 781-a*), s, Z (CYA 359*-CYA 781-a*), t, K (CYA")
156 134 M a b c d e f g h ı i j k l m n o p r s t 0 Şekil 4.24: DGGE de Urla, Şifne, Zeytindalı ve Karakoç termallerinin izole edilmeyen karışık kültürlerinin karşılaştırılması. Marker (M), a, U (CYA 359-CYA 781-a), b, Ş (CYA 359-CYA 781-a), c, Z (CYA 359-CYA 781-a), d, K (CYA 359-CYA 781-a); e, U (CYA 359-CYA 781-b), f, Ş (CYA 359-CYA 781-b), g, Z (CYA 359-CYA 781-b), h, K 4 (CYA 359-CYA 781-b); ı, U (CYA 359-CYA 781-a*), i, Ş (CYA 359-CYA 781-a*), j, Z (CYA 359-CYA 781-a*), k, K (CYA 359-CYA 781-a*); l, U (CYA 359-CYA 781-b*), m, Ş (CYA 359-CYA 781-b*), n, Z (CYA 359- CYA 781-b*), o, K (CYA 359-CYA 781-b*), p, U (CYA 359*-CYA 781-a*), r, Ş (CYA 359*- CYA 781-a*), s, Z (CYA 359*-CYA 781-a*), t, K (CYA 359*-CYA 781-a*).
157 135 DGGE den elde edilen veriler Şekil 4.19, 4.20, 4.21, 4.22, 4.23, 4.24 te gösterilmiştir. Şekillerde tek çoğal bölgeler ve bazen tekrarlı bölgeler oklar ile belirtilmektedir. Tekrarlı bölgelerde DNA tekrarları olduğu ama tek baz değişimlerinden dolayı yakın bölgelerde çıktıkları saptanmıştır. PCR-DGGE nin tek baz değişimlerin belirlenmesinde duyarlı bir yöntem olduğu kanıtlanmıştır. Siyanotoksinlerin oluşumu, kimyasal özellikleri ve toksik etkileri, büyük ölçüde çalışılmış fakat bunların fizyolojik fonksiyonları, regülasyonları ve bunların üretim sebepleri hala bilinmemektedir. Bazı varsayılarla siyanotoksin fonksiyonlarının ana hatlarını çıkarmıştır: (1) zooplankton ve yüksek hayvanlar gibi organizmaların siyanobakterilerin olduğu yerlerden otlamasının engellenmesi, (2) popülasyon yapısındaki değişimlerin neden olduğu ekolojik avantajlarla kazanç elde etmeleri, (3) hücre sinyal allelopatisinde ara buluculuk ve kemotaksi ile diğer siyanobakteriler veya diğer organizmalarla trofik ilişkiler kurmasıdır (Osswald et al., 2007a). Çeşitli immünolojik ve biyokimyasal belirleme yöntemleri çevre örneklerinde ve laboratuvarda siyanotoksinlerin tespitinde kullanıldığı kanıtlamıştır. Örneğin, ELİSA ve kolorimetrik (protein fosfataz inhibisyonu) deneyleri hepatotoksinlerden, mikrosistin ve nodularinin belirlenmesinde kullanılmaktadır. Ancak, moleküler belirleme yöntemleri, özellikle polimeraz zincir reaksiyonu (PCR) dayalı olanlar bu alanda giderek artan popülerliğe sahip oldukları kanıtlamaktadır. Ayrıca toksik siyanobakteriler için kolay, hızlı ve maliyet-etkin, güncel hızlı moleküler algılama yöntemlerinin son derece spesifik, hassas ve yüksek verimli analizler olduğu kabul edilmektedir (Pearson et al., 2008; Sivonen, 2008). Tarama yöntemlerinden ELİSA siyanotoksinlerin peptid geliştirilmiş ve duyarlılığı kanıtlanmıştır. Birçok araştırmacı mikrosistinlere karşı antikorlar üretmiştir. Örneğin, Kfir et al., (1986) Mikrosistin-LA ya karşı monoklonal antikorlar üretmiş, diğer mikrosistinler eşit verimlilikte geniş bağlanma kapasitesine sahiptir. Microcystis in bir türünün toksinlerine karşı antikor üretmiş, diğer toksik Microcystis suşları çapraz reaksiyona girmiş fakat toksinleri içeren peptid diğer cinslerin suşları ile reaksiyona girmemektedir. Ancak, sonra

 mikrosistin ELİSA tekniklerini kullanılarak çok sayıda çapraz reaksiyonları değerlendirmiş, reaktivite ve akut toksisitesite arasında zayıf bir korelasyon bulmuştur.")
158 136 çalışmalar örneğin değişken çapraz reaktivitesini göstermiştir, örneğin Mikrosistin-LR karşı antikorlar oluşturmuş ve (Mikrosistin-RR) için iyi (Mikrosistin-LA) için ise kötü olan çapraz reaksiyonlar arasında farklılık bulmuştur. Ancak, bu işlem çevre örneklerinde başarıyla mikrosistin içeriğini belirlemek için kullanılmıştır. Başka bir gelişmede, Mcdermott et al., (1995) Mikrosistin-LR karşı üretilen antikorlar Mikrosistin-RR ile yüksek çapraz reaksiyon göstermiş ve orada iyi bir anlaşma olduğunu ELİSA ve HPLC sonuçları ile doğrulanmıştır. An and Carmichael, (1994) mikrosistin ELİSA tekniklerini kullanılarak çok sayıda çapraz reaksiyonları değerlendirmiş, reaktivite ve akut toksisitesite arasında zayıf bir korelasyon bulmuştur. Bu ELİSA teknikleri bazı Mikrosistin türevlerinin konsantrasyonu azalttığı göstermiştir (Msagatia et al., 2006). Kullandığımız Envirollogix marka ELİSA kiti Mikrotoksin-LR, Mikrotoksin-LA, Microcystin-RR, Microcystin-YR ve Nodularin ile çapraz reaksiyona girmektedir. Sonuçları 0,16 ppb nin altında çıkmıştır (Çizelge 4.6, Şekil 4.25). A B Şekil 4.25 : ELİSA platenin A. Durdurma çözeltisi eklenmeden önce B. Durdurma çözeltisi eklendikten sonraki görüntüsü
159 137 Çizelge 4.6: ELİSA sonuçları Kalibratör % Bo 0,16 ppb 88,6689 0,6 ppb 48, ,5 ppb 15,38227 U sal 97,47 ZD 89,92 KY 90,59 Ş 92,39 Bal pse 88,79 U osc 89,49 Bal ip 94,58 KK 88,09 U pse 89,31 U ip 89,92 Siyanobakterilerin hepatotoksik olanları ve toksik olmayan türlerinde farklı mikrosistin sentetaz (mcy) genlerinin içeriği bulunmaktadır (Oberholster and Botha, 2007, Tillett et al., 2000, Rouhıaınen et al., 2004). Bu nedenle izolatların stasyonel fazda DNA ları elde edilerek toksin gen bölgeleri için seçilen primer çiftleri ile PCR yapılmış ve toksin gen bölgelerinin var olduğu görülmüştür (Şekil 4.26, 4.27, 4.28). Sıcak su kaynakları dünya çevresinde kolay bulunmaktadır, örneğin Suudi Arabistan da, halk ve turistler için eğlence ve bazı yerel komüniteler bu suları yemeklerinde de kullanmaktadır. Sıcak su içinde bulunan siyanobakteri kümeleriyle ve onların toksinlerinin kontamine ettiği sularla deri teması ve kazara yutmak, banyo yapmak veya eğlenmek sağlık riskine neden olabilmektedir. Kısacası sıcak su kaynaklarında bulunan siyanobakteriyel kümelerle, banyo ve su eğlencesi yapan insanların derilerinde ve gözlerinde tahrişe neden olmaktadır (Mohamed, 2008) yılında Brezilya nın Caruaru şehrinde yüksek mikrosistin konsantrasyonuna maruz kalan 60 hemodiyaliz hastasının ölmesiyle büyük bir trajedi yaşanmıştır. Bu olaydan sonra Brezilya Sağlık Bakanlığı tarafından su kaynaklarında siyanotoksinlerin izlenmesi için bir kılavuz hazırlanmıştır ve bu kılavuzda MC-LR için limit değer 1 µgl -1 olmasına karar verilmiştir. Su içinde
. Şekil 4.")
; 1. U ip; 2. Bal pse; 3.")
160 138 MC-LR düşük konsantrasyonları bile büyük endişeye neden olmasında akut toksisite ve bu doğal bileşiklerin sublethal toksisitesi önemli rol oynamaktadır (Anjos et al., 2006). Şekil 4.26: Saksitoksin gen bölgelerinin görüntüsü; Marker (M); 1. Şif; 2. Bal pse; 3. ZD; 4. U osc; 5. U ip; 6. U sal; 7. KK; 8. Bal ip; 9. KY; 10. U sal Şekil 4.27: Mikrosistin A gen bölgelerinin görüntüsü, Marker (M); 1. U ip; 2. Bal pse; 3.U pse; 4. Şif; 5. ZD; 6. Usal; 7. KK; 8. Uosc; 9. KY; 10. Bal ip

161 139 Şekil 4. 28: Nodularin gen bölgesinin görüntüsü, M; 1. KY; 2. ZD; 3.U sal; 4. U osc; 5. Bal ip; 6. Uip; 7. Şif; 8. Bal pse; 9. U pse; 10. KK Şekil 4.26, 4.27, 4.28 de türlerin toksin gen bölgeleri içerdiği saptanmıştır. Toksin gen bölgelerini içeren türlerin ELİSA sonuçları ile karşılaştırıldığında hastalık yapıcı değerlerde olmadığı belirlenmiştir. Toksin gen bölgeleri toplam gen bölgelerininden seçilen kısımlar olduğundan diğer gen bölgelerini de içerip içermedikleri belirlenmelidir. Toksin gen bölgelerini uyaracak operonlar aktif hale geçirilmediği için toksik etki göstermediği düşünülmektedir. Siyanobakterilerin bu toksinleri ani çoğalamalarında ve uygunsuz koşullarda hayatta kalmak için salgıladığı düşünüldüğünde üretim koşulları değiştirilerek uyarıcaların değişimlerinin gen bölgeleri üzerine etkileri izlenmeli ve karşılaştırılmalıdır. Sonuç olarak İzmir ilindeki termal alanlardan izole edilen ve moleküler olarak tanımlanan türlerin toksin gen bölgelerinin varlığı belirlenmiş olsa bile toksisite göstermedikleri belirlenmiştir. Elde edilen bu bilgiler ışığında bu türlerin diğer çalışmalarda da kullanılması için kültür koleksiyonunda toplanarak devamlılığının sağlanması düşünülmektedir. Dizi analizi sonuçları uluslarası bir dizi veri bankasına kayıt ettirilmiş ve İzmir ilindeki termal alanlarda yaşayan siyanobakteri türlerin dizileri kayıt altına alınması sağlanmıştır.
İÇME SULARINDA YÜKSEK HASSASİYETTE SİYANOTOKSİN ANALİZİ
 İÇME SULARINDA YÜKSEK HASSASİYETTE SİYANOTOKSİN ANALİZİ Cevdet Demir, Habibe Ajderoğlu Uludağ Üniversitesi, Fen Edebiyat Fakültesi, Kimya Bölümü, 16059 BURSA BUSKİ Dobruca İçme Suyu Arıtma Tesisi, İçme
İÇME SULARINDA YÜKSEK HASSASİYETTE SİYANOTOKSİN ANALİZİ Cevdet Demir, Habibe Ajderoğlu Uludağ Üniversitesi, Fen Edebiyat Fakültesi, Kimya Bölümü, 16059 BURSA BUSKİ Dobruca İçme Suyu Arıtma Tesisi, İçme
POLİMERAZ ZİNCİR REAKSİYONU (PZR-PCR) VE RESTRİKSİYON PARÇA UZUNLUĞU POLİMORFİZMİ (RFLP)
 VE RESTRİKSİYON PARÇA UZUNLUĞU POLİMORFİZMİ (RFLP)") Deney: M 1 POLİMERAZ ZİNCİR REAKSİYONU (PZR-PCR) VE RESTRİKSİYON PARÇA UZUNLUĞU POLİMORFİZMİ (RFLP) a) PCR yöntemi uygulaması b) RPLF sonuçları değerlendirilmesi I. Araç ve Gereç dntp (deoksi Nükleotid
Deney: M 1 POLİMERAZ ZİNCİR REAKSİYONU (PZR-PCR) VE RESTRİKSİYON PARÇA UZUNLUĞU POLİMORFİZMİ (RFLP) a) PCR yöntemi uygulaması b) RPLF sonuçları değerlendirilmesi I. Araç ve Gereç dntp (deoksi Nükleotid
CYANOBACTERIOPHYTA (Mavi-Yeşil Algler)
") CYANOBACTERIOPHYTA (Mavi-Yeşil Algler) Bu gruba giren bitkiler, tek hücreli veya koloni halinda yaşayan, bazı üyeleri ipliksi yapıda olan mavi renkli alglerdir. Bir kısmı bakteri kadar küçüktür. Bir hücrede
CYANOBACTERIOPHYTA (Mavi-Yeşil Algler) Bu gruba giren bitkiler, tek hücreli veya koloni halinda yaşayan, bazı üyeleri ipliksi yapıda olan mavi renkli alglerdir. Bir kısmı bakteri kadar küçüktür. Bir hücrede
BAKTERİLERİN GENETİK KARAKTERLERİ
 BAKTERİLERİN GENETİK KARAKTERLERİ GENETİK MATERYALLER VE YAPILARI HER HÜCREDE Genetik bilgilerin kodlandığı bir DNA genomu bulunur Bu genetik bilgiler mrna ve ribozomlar aracılığı ile proteinlere dönüştürülür
BAKTERİLERİN GENETİK KARAKTERLERİ GENETİK MATERYALLER VE YAPILARI HER HÜCREDE Genetik bilgilerin kodlandığı bir DNA genomu bulunur Bu genetik bilgiler mrna ve ribozomlar aracılığı ile proteinlere dönüştürülür
MOLEKÜLER BİYOLOJİ LABORATUVARI
 MOLEKÜLER 2014-2015 BİYOLOJİ LABORATUVARI GÜZ DÖNEMİ MOLEKÜLER BİYOLOJİ LABORATUVARI 7.HAFTA DERS NOTLARI GAZİ ÜNİVERSİTESİ FEN FAKÜLTESİ BİYOLOJİ BÖLÜMÜ Sayfa 1 / 6 1. RFLP (RESTRİKSİYON PARÇA UZUNLUK
MOLEKÜLER 2014-2015 BİYOLOJİ LABORATUVARI GÜZ DÖNEMİ MOLEKÜLER BİYOLOJİ LABORATUVARI 7.HAFTA DERS NOTLARI GAZİ ÜNİVERSİTESİ FEN FAKÜLTESİ BİYOLOJİ BÖLÜMÜ Sayfa 1 / 6 1. RFLP (RESTRİKSİYON PARÇA UZUNLUK
Siyanotoksinler ve Biyosentezlerinden Sorumlu Genlerin Düzenlenmesi
 ISSN: 2146-8168 Sayı: 11, Yıl: 2015, Sayfa: 54-69 http://bilader.gop.edu.tr Dergiye Geliş Tarihi: 05.08.2015 Yayına Kabul Tarihi: 10.09.2015 Baş Editör: Bilge Hilal ÇADIRCI Alan Editörü: Kubilay YILDIRIM
ISSN: 2146-8168 Sayı: 11, Yıl: 2015, Sayfa: 54-69 http://bilader.gop.edu.tr Dergiye Geliş Tarihi: 05.08.2015 Yayına Kabul Tarihi: 10.09.2015 Baş Editör: Bilge Hilal ÇADIRCI Alan Editörü: Kubilay YILDIRIM
TRANSLASYON ve PROTEİNLER
 TRANSLASYON ve PROTEİNLER Prof. Dr. Sacide PEHLİVAN 13 Aralık 2016 mrna daki baz sırasının kullanılarak amino asitlerin doğru sıra ile proteini oluşturmasını kapsayan olayların tümüne Translasyon veya
TRANSLASYON ve PROTEİNLER Prof. Dr. Sacide PEHLİVAN 13 Aralık 2016 mrna daki baz sırasının kullanılarak amino asitlerin doğru sıra ile proteini oluşturmasını kapsayan olayların tümüne Translasyon veya
Akvaryum veya küçük havuzlarda amonyağın daha az zehirli olan nitrit ve nitrata dönüştürülmesi için gerekli olan bakteri populasyonunu (nitrifikasyon
 Azotlu bileşikler Ticari balık havuzlarında iyonize olmuş veya iyonize olmamış amonyağın konsantrasyonlarını azaltmak için pratik bir yöntem yoktur. Balık havuzlarında stoklama ve yemleme oranlarının azaltılması
Azotlu bileşikler Ticari balık havuzlarında iyonize olmuş veya iyonize olmamış amonyağın konsantrasyonlarını azaltmak için pratik bir yöntem yoktur. Balık havuzlarında stoklama ve yemleme oranlarının azaltılması
BİTKİ BESLEME DERS NOTLARI
 BİTKİ BESLEME DERS NOTLARI Dr. Metin AYDIN KONYA 2011 BİTKİ BESİN ELEMENTLERİNİN GÖREVLERİ, ALINIŞ FORMLARI ve KAYNAKLARI Besin Elementi Bitkideki Görevi Alınış Formu Kaynakları Karbon (C) Karbonhidratların
BİTKİ BESLEME DERS NOTLARI Dr. Metin AYDIN KONYA 2011 BİTKİ BESİN ELEMENTLERİNİN GÖREVLERİ, ALINIŞ FORMLARI ve KAYNAKLARI Besin Elementi Bitkideki Görevi Alınış Formu Kaynakları Karbon (C) Karbonhidratların
15- RADYASYONUN NÜKLEİK ASİTLER VE PROTEİNLERE ETKİLERİ
 15- RADYASYONUN NÜKLEİK ASİTLER VE PROTEİNLERE ETKİLERİ İyonlaştırıcı radyasyonların biyomoleküllere örneğin nükleik asitler ve proteinlere olan etkisi hakkında yeterli bilgi yoktur. Ancak, nükleik asitlerden
15- RADYASYONUN NÜKLEİK ASİTLER VE PROTEİNLERE ETKİLERİ İyonlaştırıcı radyasyonların biyomoleküllere örneğin nükleik asitler ve proteinlere olan etkisi hakkında yeterli bilgi yoktur. Ancak, nükleik asitlerden
Çizelge 2.6. Farklı ph ve su sıcaklığı değerlerinde amonyak düzeyi (toplam amonyağın yüzdesi olarak) (Boyd 2008a)
 (Boyd 2008a)") - Azotlu bileşikler Su ürünleri yetiştiricilik sistemlerinde oksijen gereksinimi karşılandığı takdirde üretimi sınırlayan ikinci faktör azotlu bileşiklerin birikimidir. Ana azotlu bileşikler; azot gazı
- Azotlu bileşikler Su ürünleri yetiştiricilik sistemlerinde oksijen gereksinimi karşılandığı takdirde üretimi sınırlayan ikinci faktör azotlu bileşiklerin birikimidir. Ana azotlu bileşikler; azot gazı
III-Hayatın Oluşturan Kimyasal Birimler
 III-Hayatın Oluşturan Kimyasal Birimler MBG 111 BİYOLOJİ I 3.1.Karbon:Biyolojik Moleküllerin İskeleti *Karbon bütün biyolojik moleküllerin omurgasıdır, çünkü dört kovalent bağ yapabilir ve uzun zincirler
III-Hayatın Oluşturan Kimyasal Birimler MBG 111 BİYOLOJİ I 3.1.Karbon:Biyolojik Moleküllerin İskeleti *Karbon bütün biyolojik moleküllerin omurgasıdır, çünkü dört kovalent bağ yapabilir ve uzun zincirler
2. Histon olmayan kromozomal proteinler
 12. Hafta: Nükleik Asitler: Nükleik asitlerin yapısal üniteleri, nükleozitler, nükleotidler, inorganik fosfat, nükleotidlerin fonksiyonları, nükleik asitler, polinükleotidler, DNA nın primer ve sekonder
12. Hafta: Nükleik Asitler: Nükleik asitlerin yapısal üniteleri, nükleozitler, nükleotidler, inorganik fosfat, nükleotidlerin fonksiyonları, nükleik asitler, polinükleotidler, DNA nın primer ve sekonder
REKOMBİNANT DNA TEKNOLOJİSİ. Araş. Gör. Dr. Öğünç MERAL
 Araş. Gör. Dr. Öğünç MERAL 1960 lardan bu yana genetik ve moleküler biyolojideki kavrayışımızın hızla artması, biyoteknolojide heyecan verici buluşlar ve uygulamalara yol açtı. DNA yapısı ve fonksiyonlarının
Araş. Gör. Dr. Öğünç MERAL 1960 lardan bu yana genetik ve moleküler biyolojideki kavrayışımızın hızla artması, biyoteknolojide heyecan verici buluşlar ve uygulamalara yol açtı. DNA yapısı ve fonksiyonlarının
Agaroz jel elektroforezi
 MOLEKÜLER TEKNİKLER Dr. Naşit İĞCİ Nevşehir Hacı Bektaş Veli Üniversitesi Moleküler Biyoloji ve Genetik Bölümü 4. Sınıf (2017-2018 Bahar) 2. NOT Agaroz jel elektroforezi PAGE daha çok proteinlerin ve küçük
MOLEKÜLER TEKNİKLER Dr. Naşit İĞCİ Nevşehir Hacı Bektaş Veli Üniversitesi Moleküler Biyoloji ve Genetik Bölümü 4. Sınıf (2017-2018 Bahar) 2. NOT Agaroz jel elektroforezi PAGE daha çok proteinlerin ve küçük
BİYOİNORGANİK KİMYA 5. HAFTA
 BİYOİNORGANİK KİMYA 5. HAFTA ESER ELEMENTLER İnsan vücudunda en yüksek oranda bulunan element oksijendir. İkincisi ise karbondur. İnsan vücudunun kütlesinin %99 u sadece 6 elementten meydana gelir. Bunlar:
BİYOİNORGANİK KİMYA 5. HAFTA ESER ELEMENTLER İnsan vücudunda en yüksek oranda bulunan element oksijendir. İkincisi ise karbondur. İnsan vücudunun kütlesinin %99 u sadece 6 elementten meydana gelir. Bunlar:
TRANSLASYON VE DÜZENLENMESİ
 TRANSLASYON VE DÜZENLENMESİ TRANSLASYON Translasyonda nükleik asit kullanılır fakat son ürün bir nükleik asit değil proteindir. Translasyon mekanizması 4 ana bileşenden oluşmaktadır: 1. mrnalar 2. trnalar
TRANSLASYON VE DÜZENLENMESİ TRANSLASYON Translasyonda nükleik asit kullanılır fakat son ürün bir nükleik asit değil proteindir. Translasyon mekanizması 4 ana bileşenden oluşmaktadır: 1. mrnalar 2. trnalar
2006 ÖSS BİYOLOJİ SORULARI VE CEVAPLARI
 2006 ÖSS BİYOLOJİ SORULARI VE CEVAPLARI 1. BÖLÜM 1. I. Adaptasyon II. Mutasyon III. Kalıtsal varyasyon Bir populasyondaki bireyler, yukarıdakilerden hangilerini "doğal seçilim ile kazanır? D) I veii E)
2006 ÖSS BİYOLOJİ SORULARI VE CEVAPLARI 1. BÖLÜM 1. I. Adaptasyon II. Mutasyon III. Kalıtsal varyasyon Bir populasyondaki bireyler, yukarıdakilerden hangilerini "doğal seçilim ile kazanır? D) I veii E)
TEST 1. Hücre Solunumu. 4. Aşağıda verilen moleküllerden hangisi oksijenli solunumda substrat olarak kullanılamaz? A) Glikoz B) Mineral C) Yağ asidi
 Glikoz B) Mineral C) Yağ asidi") 1. Termometre Çimlenen bezelye tohumlar Termos Çimlenen bezelye tohumları oksijenli solunum yaptığına göre yukarıdaki düzenekle ilgili, I. Termostaki oksijen miktarı azalır. II. Termometredeki sıcaklık
1. Termometre Çimlenen bezelye tohumlar Termos Çimlenen bezelye tohumları oksijenli solunum yaptığına göre yukarıdaki düzenekle ilgili, I. Termostaki oksijen miktarı azalır. II. Termometredeki sıcaklık
MOLEKÜLER BİYOLOJİ DOÇ. DR. MEHMET KARACA (5. BÖLÜM)
") MOLEKÜLER BİYOLOJİ DOÇ. DR. MEHMET KARACA (5. BÖLÜM) TRANSKRİPSİYONU (ÖKARYOTİK) STOPLAZMA DNA Transkripsiyon hnrna RNA nın işlenmesi mrna G AAA Eksport G AAA NÜKLEUS TRANSKRİPSİYONU (PROKARYOTİK) Stoplazma
MOLEKÜLER BİYOLOJİ DOÇ. DR. MEHMET KARACA (5. BÖLÜM) TRANSKRİPSİYONU (ÖKARYOTİK) STOPLAZMA DNA Transkripsiyon hnrna RNA nın işlenmesi mrna G AAA Eksport G AAA NÜKLEUS TRANSKRİPSİYONU (PROKARYOTİK) Stoplazma
Replikasyon, Transkripsiyon ve Translasyon. Yrd. Doç. Dr. Osman İBİŞ
 Replikasyon, Transkripsiyon ve Translasyon Yrd. Doç. Dr. Osman İBİŞ DNA replikasyonu DNA nın replikasyonu, DNA molekülünün, sakladığı genetik bilgilerin sonraki nesillere aktarılması için kendi kopyasını
Replikasyon, Transkripsiyon ve Translasyon Yrd. Doç. Dr. Osman İBİŞ DNA replikasyonu DNA nın replikasyonu, DNA molekülünün, sakladığı genetik bilgilerin sonraki nesillere aktarılması için kendi kopyasını
Örnek : 3- Bileşiklerin Özellikleri :
 Bileşikler : Günümüzde bilinen 117 element olmasına rağmen (92 tanesi doğada bulunur) bu elementler farklı sayıda ve şekilde birleşerek ve etkileşerek farklı kimyasal özelliklere sahip milyonlarca yani
Bileşikler : Günümüzde bilinen 117 element olmasına rağmen (92 tanesi doğada bulunur) bu elementler farklı sayıda ve şekilde birleşerek ve etkileşerek farklı kimyasal özelliklere sahip milyonlarca yani
Canlıların yapısına en fazla oranda katılan organik molekül çeşididir. Deri, saç, tırnak, boynuz gibi oluşumların temel maddesi proteinlerdir.
 Canlıların yapısına en fazla oranda katılan organik molekül çeşididir. Deri, saç, tırnak, boynuz gibi oluşumların temel maddesi proteinlerdir. Proteinlerin yapısında; Karbon ( C ) Hidrojen ( H ) Oksijen
Canlıların yapısına en fazla oranda katılan organik molekül çeşididir. Deri, saç, tırnak, boynuz gibi oluşumların temel maddesi proteinlerdir. Proteinlerin yapısında; Karbon ( C ) Hidrojen ( H ) Oksijen
Çevre Biyolojisi
 Çevre Biyolojisi 115-02 Bölüm 03-02 Temel Ekolojik Kavramlar ve Süreçler Çağatay Tavşanoğlu 2017-2018 Güz Biyojeokimyasal döngüler ve madde geri kazanımı Ekosistem boyunca enerji akışı tek yönlü bir süreçtir
Çevre Biyolojisi 115-02 Bölüm 03-02 Temel Ekolojik Kavramlar ve Süreçler Çağatay Tavşanoğlu 2017-2018 Güz Biyojeokimyasal döngüler ve madde geri kazanımı Ekosistem boyunca enerji akışı tek yönlü bir süreçtir
BİTKİ BESİN ELEMENTLERİ BİTKİ BESİN ELEMENTLERİ, TANIMI VE SINIFLANDIRILMASI
 BİTKİ BESİN ELEMENTLERİ BİTKİ BESİN ELEMENTLERİ, TANIMI VE SINIFLANDIRILMASI Bitkiler geliştikleri ortamdan toprak altı ve toprak üstü organlarıyla çok sayıda element (74) alır. Ancak bu elementlerin çok
BİTKİ BESİN ELEMENTLERİ BİTKİ BESİN ELEMENTLERİ, TANIMI VE SINIFLANDIRILMASI Bitkiler geliştikleri ortamdan toprak altı ve toprak üstü organlarıyla çok sayıda element (74) alır. Ancak bu elementlerin çok
6. BÖLÜM MİKROBİYAL METABOLİZMA
 6. BÖLÜM MİKROBİYAL METABOLİZMA 1 METABOLİZMA Hücrede meydana gelen tüm reaksiyonlara denir Anabolizma: Basit moleküllerden kompleks moleküllerin sentezlendiği enerji gerektiren reaksiyonlardır X+Y+ENERJİ
6. BÖLÜM MİKROBİYAL METABOLİZMA 1 METABOLİZMA Hücrede meydana gelen tüm reaksiyonlara denir Anabolizma: Basit moleküllerden kompleks moleküllerin sentezlendiği enerji gerektiren reaksiyonlardır X+Y+ENERJİ
LABORATUVAR 4: ÖKARYOTİK HÜCRELER
 LABORATUVAR 4: ÖKARYOTİK HÜCRELER Ökaryotik hücreler, sahip oldukları hücre iskeleti, nüklear membran ve organelleri içeren bölümleri ile prokaryot hücrelerden ayırt edilebilmektedir. Kimyasal analizler
LABORATUVAR 4: ÖKARYOTİK HÜCRELER Ökaryotik hücreler, sahip oldukları hücre iskeleti, nüklear membran ve organelleri içeren bölümleri ile prokaryot hücrelerden ayırt edilebilmektedir. Kimyasal analizler
Zirai-Endüstriyel Atıksularda Siyanobakterilerden Biyokütle Eldesi ve Gübre Olarak Kullanılması
 Zirai-Endüstriyel Atıksularda Siyanobakterilerden Biyokütle Eldesi ve Gübre Olarak Kullanılması Dr. N. Oya SAN Bilkent Üniversitesi, UNAM, Malzeme Bilimi ve Nanoteknoloji Enstitüsü Doktora Sonrası Araştırmacı
Zirai-Endüstriyel Atıksularda Siyanobakterilerden Biyokütle Eldesi ve Gübre Olarak Kullanılması Dr. N. Oya SAN Bilkent Üniversitesi, UNAM, Malzeme Bilimi ve Nanoteknoloji Enstitüsü Doktora Sonrası Araştırmacı
İçme Sularının Dezenfeksiyonunda Çinko Oksit Nanomateryalinin Kullanımı
 İçme Sularının Dezenfeksiyonunda Çinko Oksit Nanomateryalinin Kullanımı F. Elçin Erkurt, Behzat Balcı, E. Su Turan Çukurova Üniversitesi Çevre Mühendisliği Bölümü Giriş Su, tüm canlılar için en önemli
İçme Sularının Dezenfeksiyonunda Çinko Oksit Nanomateryalinin Kullanımı F. Elçin Erkurt, Behzat Balcı, E. Su Turan Çukurova Üniversitesi Çevre Mühendisliği Bölümü Giriş Su, tüm canlılar için en önemli
Organik Bileşikler. Karbonhidratlar. Organik Bileşikler YGS Biyoloji 1
 Organik Bileşikler YGS Biyoloji 1 Hazırladığımız bu yazıda; organik bileşikler ve organik bileşiklerin yapısını, canlılarda bulunan organik bileşikleri ve bunların görevlerini, kullanım alanlarını, canlılar
Organik Bileşikler YGS Biyoloji 1 Hazırladığımız bu yazıda; organik bileşikler ve organik bileşiklerin yapısını, canlılarda bulunan organik bileşikleri ve bunların görevlerini, kullanım alanlarını, canlılar
7. PROKARYOTLARDA GEN İFADESİNİN DÜZENLENMESİ
 7. PROKARYOTLARDA GEN İFADESİNİN DÜZENLENMESİ Başlıklar 1. Prokaryotlar gen ifadesini çevre koşullarına göre düzenler 2. E. Coli de laktoz metabolizması 3. Lac operonu negatif kontrol 4. CAP pozitif kontrol
7. PROKARYOTLARDA GEN İFADESİNİN DÜZENLENMESİ Başlıklar 1. Prokaryotlar gen ifadesini çevre koşullarına göre düzenler 2. E. Coli de laktoz metabolizması 3. Lac operonu negatif kontrol 4. CAP pozitif kontrol
olmak üzere 2 gruba ayrılırlar.
 CANLI ALEMLERİ *Canlı alemleri: Canlılar filogenetik sınıflandırmaya göre; bakteriler, arkebakteriler, protista, fungi, bitki ve hayvanlar olmak üzere 6 Aleme ayrılır. **Prokaryot canlılar: Çekirdeksiz
CANLI ALEMLERİ *Canlı alemleri: Canlılar filogenetik sınıflandırmaya göre; bakteriler, arkebakteriler, protista, fungi, bitki ve hayvanlar olmak üzere 6 Aleme ayrılır. **Prokaryot canlılar: Çekirdeksiz
POYRAZ TIBBİ CİHAZLAR EDVOTEK
 POYRAZ TIBBİ CİHAZLAR EDVOTEK EDVOTEK VİZYON Edvotek, bir çok disiplini bir araya getirerek karmaşık gibi görünen birçok bilimin temellerini anlatarak «Nasıl bilim adamı yetiştiririz?» sorusuna karşılık
POYRAZ TIBBİ CİHAZLAR EDVOTEK EDVOTEK VİZYON Edvotek, bir çok disiplini bir araya getirerek karmaşık gibi görünen birçok bilimin temellerini anlatarak «Nasıl bilim adamı yetiştiririz?» sorusuna karşılık
TÜBİTAK BİDEB LİSE ÖĞRETMENLERİ FİZİK, KİMYA, BİYOLOJİ, MATEMATİK- PROJE DANIŞMANLIĞI EĞİTİMİ ÇALIŞTAYI LİSE3 (Çalıştay 2013) BİYOLOJİ GRUP TUHAF
 BİYOLOJİ GRUP TUHAF") TÜBİTAK BİDEB LİSE ÖĞRETMENLERİ FİZİK, KİMYA, BİYOLOJİ, MATEMATİK- PROJE DANIŞMANLIĞI EĞİTİMİ ÇALIŞTAYI LİSE3 (Çalıştay 2013) BİYOLOJİ GRUP TUHAF PROJE ÖNERİSİ ADI TUHAF MATERYALLERDEN İZOLE EDİLEN DNA
TÜBİTAK BİDEB LİSE ÖĞRETMENLERİ FİZİK, KİMYA, BİYOLOJİ, MATEMATİK- PROJE DANIŞMANLIĞI EĞİTİMİ ÇALIŞTAYI LİSE3 (Çalıştay 2013) BİYOLOJİ GRUP TUHAF PROJE ÖNERİSİ ADI TUHAF MATERYALLERDEN İZOLE EDİLEN DNA
Stres Koşulları ve Bitkilerin Tepkisi
 Stres Koşulları ve Bitkilerin Tepkisi Stres nedir? Olumsuz koşullara karşı canlıların vermiş oldukları tepkiye stres denir. Olumsuz çevre koşulları bitkilerde strese neden olur. «Biyolojik Stres»: Yetişme
Stres Koşulları ve Bitkilerin Tepkisi Stres nedir? Olumsuz koşullara karşı canlıların vermiş oldukları tepkiye stres denir. Olumsuz çevre koşulları bitkilerde strese neden olur. «Biyolojik Stres»: Yetişme
Gram (+)Bakterilerde Duvar Yapısı Gram (-) Bakterilerde Duvar Yapısı Lipopolisakkaritin Önemi
Bakterilerde Duvar Yapısı Gram (-) Bakterilerde Duvar Yapısı Lipopolisakkaritin Önemi") Gram (+)Bakterilerde Duvar Yapısı Çoğunluğu peptidoglikan yapıdır. Bunun yanında teikoik asitte içerirler. Bu yapı gliserol veya Ribitolün PO4 gruplarına bağlanmasıyla oluşur. Teikoik asitler peptidoglikan
Gram (+)Bakterilerde Duvar Yapısı Çoğunluğu peptidoglikan yapıdır. Bunun yanında teikoik asitte içerirler. Bu yapı gliserol veya Ribitolün PO4 gruplarına bağlanmasıyla oluşur. Teikoik asitler peptidoglikan
Algisidal Deneylerde Kullanılan Mikroorganizmaların Revizyonu Amacıyla Havuzlarda Bulunan Yaygın Mikroalglerin Belirlenmesi
 Algisidal Deneylerde Kullanılan Mikroorganizmaların Revizyonu Amacıyla Havuzlarda Bulunan Yaygın Mikroalglerin Belirlenmesi Burçin Karabey 1, İnci Tüney Kızılkaya 2, Ataç Uzel 1, Güven Özdemir 1 1 Ege
Algisidal Deneylerde Kullanılan Mikroorganizmaların Revizyonu Amacıyla Havuzlarda Bulunan Yaygın Mikroalglerin Belirlenmesi Burçin Karabey 1, İnci Tüney Kızılkaya 2, Ataç Uzel 1, Güven Özdemir 1 1 Ege
7. PROKARYOTLARDA GEN İFADESİNİN DÜZENLENMESİ
 7. PROKARYOTLARDA GEN İFADESİNİN DÜZENLENMESİ Başlıklar 1. Prokaryotlar gen ifadesini çevre koşullarına göre düzenler 2. E. Coli de laktoz metabolizması 3. Lac operonu negatif kontrol 4. CAP pozitif kontrol
7. PROKARYOTLARDA GEN İFADESİNİN DÜZENLENMESİ Başlıklar 1. Prokaryotlar gen ifadesini çevre koşullarına göre düzenler 2. E. Coli de laktoz metabolizması 3. Lac operonu negatif kontrol 4. CAP pozitif kontrol
CANLILARIN YAPISINDA BULUNAN TEMEL BİLEŞENLER
 CANLILARIN YAPISINDA BULUNAN TEMEL BİLEŞENLER Canlıların yapısında bulunan moleküller yapısına göre 2 ye ayrılır: I. İnorganik Bileşikler: Bir canlı vücudunda sentezlenemeyen, dışardan hazır olarak aldığı
CANLILARIN YAPISINDA BULUNAN TEMEL BİLEŞENLER Canlıların yapısında bulunan moleküller yapısına göre 2 ye ayrılır: I. İnorganik Bileşikler: Bir canlı vücudunda sentezlenemeyen, dışardan hazır olarak aldığı
YAZILIYA HAZIRLIK SORULARI. 11. Sınıf 1 CANLILARDA ENERJİ DÖNÜŞÜMLERİ
 YAZILIYA HAZIRLIK SORULARI 11. Sınıf 1 CANLILARDA ENERJİ DÖNÜŞÜMLERİ Fotosentez ile ışık enerjisi kimyasal bağ enerjisine dönüştürülür. Kloroplastsız hücreler fotosentez yapamaz. DOĞRU YANLIŞ SORULARI
YAZILIYA HAZIRLIK SORULARI 11. Sınıf 1 CANLILARDA ENERJİ DÖNÜŞÜMLERİ Fotosentez ile ışık enerjisi kimyasal bağ enerjisine dönüştürülür. Kloroplastsız hücreler fotosentez yapamaz. DOĞRU YANLIŞ SORULARI
YAZILIYA HAZIRLIK SORULARI. 9. Sınıf
 YAZILIYA HAZIRLIK SORULARI 9. Sınıf DOĞRU YANLIŞ SORULARI Nitel gözlemlerin güvenilirliği nicel gözlemlerden fazladır. Ökaryot hücrelerde kalıtım materyali çekirdek içinde bulunur. Ototrof beslenen canlılar
YAZILIYA HAZIRLIK SORULARI 9. Sınıf DOĞRU YANLIŞ SORULARI Nitel gözlemlerin güvenilirliği nicel gözlemlerden fazladır. Ökaryot hücrelerde kalıtım materyali çekirdek içinde bulunur. Ototrof beslenen canlılar
KİMYA-IV. Yrd. Doç. Dr. Yakup Güneş
 KİMYA-IV Yrd. Doç. Dr. Yakup Güneş Organik Kimyaya Giriş Kimyasal bileşikler, eski zamanlarda, elde edildikleri kaynaklara bağlı olarak Anorganik ve Organik olmak üzere, iki sınıf altında toplanmışlardır.
KİMYA-IV Yrd. Doç. Dr. Yakup Güneş Organik Kimyaya Giriş Kimyasal bileşikler, eski zamanlarda, elde edildikleri kaynaklara bağlı olarak Anorganik ve Organik olmak üzere, iki sınıf altında toplanmışlardır.
Hücrelerde gerçekleşen yapım, yıkım ve dönüşüm olaylarının bütününe metabolizma denir.
 METABOLİZMA ve ENZİMLER METABOLİZMA Hücrelerde gerçekleşen yapım, yıkım ve dönüşüm olaylarının bütününe metabolizma denir. A. ÖZÜMLEME (ANABOLİZMA) Metabolizmanın yapım reaksiyonlarıdır. Bu tür olaylara
METABOLİZMA ve ENZİMLER METABOLİZMA Hücrelerde gerçekleşen yapım, yıkım ve dönüşüm olaylarının bütününe metabolizma denir. A. ÖZÜMLEME (ANABOLİZMA) Metabolizmanın yapım reaksiyonlarıdır. Bu tür olaylara
Biochemistry Chapter 4: Biomolecules. Hikmet Geçkil, Professor Department of Molecular Biology and Genetics Inonu University
 Biochemistry Chapter 4: Biomolecules, Professor Department of Molecular Biology and Genetics Inonu University Biochemistry/Hikmet Geckil Chapter 4: Biomolecules 2 BİYOMOLEKÜLLER Bilim adamları hücreyi
Biochemistry Chapter 4: Biomolecules, Professor Department of Molecular Biology and Genetics Inonu University Biochemistry/Hikmet Geckil Chapter 4: Biomolecules 2 BİYOMOLEKÜLLER Bilim adamları hücreyi
Ders Kodu Ders Adı Ders Türü AKTS Hafta Teorik
 Önlisans - Sağlık Hizmetleri Meslek Yüksekokulu - Tıbbi Laboratuvar Teknikleri Y : Yıl D : Dönem Ders Kodu Ders Adı Ders Türü Y D AKTS TLT137 Genel Biyoloji Zorunlu 1 1 4 Dersin Amacı Prokaryotik ve ökaryotik
Önlisans - Sağlık Hizmetleri Meslek Yüksekokulu - Tıbbi Laboratuvar Teknikleri Y : Yıl D : Dönem Ders Kodu Ders Adı Ders Türü Y D AKTS TLT137 Genel Biyoloji Zorunlu 1 1 4 Dersin Amacı Prokaryotik ve ökaryotik
PROKARYOTLARDA GEN EKSPRESYONU. ve REGÜLASYONU. (Genlerin Gen Ürünlerine Dönüşümünü Kontrol Eden Süreçler)
") PROKARYOTLARDA GEN EKSPRESYONU ve REGÜLASYONU (Genlerin Gen Ürünlerine Dönüşümünü Kontrol Eden Süreçler) Nihal EYVAZ (050559015) Şerife OKAY (050559025) Prof. Dr. Figen ERKOÇ Gazi Eğitim Fakültesi Gen
PROKARYOTLARDA GEN EKSPRESYONU ve REGÜLASYONU (Genlerin Gen Ürünlerine Dönüşümünü Kontrol Eden Süreçler) Nihal EYVAZ (050559015) Şerife OKAY (050559025) Prof. Dr. Figen ERKOÇ Gazi Eğitim Fakültesi Gen
Yetiştirme Ortamlarında Besin Maddesi Durumunun Değerlendirilmesi
 Yetiştirme Ortamlarında Besin Maddesi Durumunun Değerlendirilmesi N, P, K ve Mg un 1:5 ekstraksiyon çözeltisindeki standard değerleri Çok az Az Yeterli Fazla Çok fazla Oldukça fazla N (meq/l)
Yetiştirme Ortamlarında Besin Maddesi Durumunun Değerlendirilmesi N, P, K ve Mg un 1:5 ekstraksiyon çözeltisindeki standard değerleri Çok az Az Yeterli Fazla Çok fazla Oldukça fazla N (meq/l)
Bitkide Fosfor. Aktif alım açısından bitki tür ve çeşitleri arasında farklılıklar vardır
 Fosfor alımı ve taşınımı Kök hücreleri ve > Bitkide Fosfor ksilem özsuyunun P kapsamı > toprak çözeltisinin P kapsamı (100-1000 kat) P alımı aktif alım şeklinde gerçekleşir Aktif alım açısından bitki tür
Fosfor alımı ve taşınımı Kök hücreleri ve > Bitkide Fosfor ksilem özsuyunun P kapsamı > toprak çözeltisinin P kapsamı (100-1000 kat) P alımı aktif alım şeklinde gerçekleşir Aktif alım açısından bitki tür
MESS Entegre Geri Kazanım ve Enerji San. ve Tic. A.Ş.
 Sayfa : 1 / 12 1 ATIKLAR İÇİN NUMUNE SAKLAMA KOŞULLARI Parametre Numune Özelliği Numune Türü ICP ile Metal Tayinleri suları vb.), diğer her türlü sıvılar) Mikrodalgada (sıvı) yakılmış Minimum Numune Miktarı
Sayfa : 1 / 12 1 ATIKLAR İÇİN NUMUNE SAKLAMA KOŞULLARI Parametre Numune Özelliği Numune Türü ICP ile Metal Tayinleri suları vb.), diğer her türlü sıvılar) Mikrodalgada (sıvı) yakılmış Minimum Numune Miktarı
BİTKİ BESİN MADDELERİ (BBM)
") BİTKİ BESİN MADDELERİ (BBM) Toprak Bilgisi Dersi Prof. Dr. Günay Erpul erpul@ankara.edu.tr Işık Enerjisinin Kimyasal Enerjiye Dönüştürülmesi Fotosentez, karbon (C), oksijen (O) ve hidrojen (H) atomlarını
BİTKİ BESİN MADDELERİ (BBM) Toprak Bilgisi Dersi Prof. Dr. Günay Erpul erpul@ankara.edu.tr Işık Enerjisinin Kimyasal Enerjiye Dönüştürülmesi Fotosentez, karbon (C), oksijen (O) ve hidrojen (H) atomlarını
İÇME SULARININ DEZENFEKSİYONUNDA NANOMATEYALLERİN KULLANIMI
 İÇME SULARININ DEZENFEKSİYONUNDA NANOMATEYALLERİN KULLANIMI Behzat Balcı, F. Elçin Erkurt, E. Su Turan Çukurova Üniversitesi Mühendislik Fakültesi Çevre Mühendisliği Bölümü Giriş İçme sularında dezenfeksiyon,
İÇME SULARININ DEZENFEKSİYONUNDA NANOMATEYALLERİN KULLANIMI Behzat Balcı, F. Elçin Erkurt, E. Su Turan Çukurova Üniversitesi Mühendislik Fakültesi Çevre Mühendisliği Bölümü Giriş İçme sularında dezenfeksiyon,
Hücre içinde bilginin akışı
 Hücre içinde bilginin akışı 1 DNA Çift Zincir Heliks 2 Hücre Çekirdeği ve Çekirdek Zarının Yapısal Organizasyonu Hatırlıyor musunuz? DNA Kromatin Kromatid Kromozom RNA Protein Çekirdek Çekirdekcik Nükleotid
Hücre içinde bilginin akışı 1 DNA Çift Zincir Heliks 2 Hücre Çekirdeği ve Çekirdek Zarının Yapısal Organizasyonu Hatırlıyor musunuz? DNA Kromatin Kromatid Kromozom RNA Protein Çekirdek Çekirdekcik Nükleotid
NÜKLEİK ASİTLERİN ELEKTROFOREZİ
 T.C. FIRAT ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ BİYOLOJİ BÖLÜMÜ NÜKLEİK ASİTLERİN ELEKTROFOREZİ Yüksek Lisans Semineri Hazırlayan: Venhar ÇELİK Danışman: Yrd.Doç.Dr. Dilek Turgut-BALIK NÜKLEİK ASİTLERİN
T.C. FIRAT ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ BİYOLOJİ BÖLÜMÜ NÜKLEİK ASİTLERİN ELEKTROFOREZİ Yüksek Lisans Semineri Hazırlayan: Venhar ÇELİK Danışman: Yrd.Doç.Dr. Dilek Turgut-BALIK NÜKLEİK ASİTLERİN
KALITSAL MOLEKÜLÜN BİÇİMİ ve ORGANİZASYONU PROF. DR. SERKAN YILMAZ
 KALITSAL MOLEKÜLÜN BİÇİMİ ve ORGANİZASYONU PROF. DR. SERKAN YILMAZ Değişik canlı gruplarında kalıtsal molekülün çeşidi, sayısı, biçimi ve organizasyonu bakımından farklılıklar bulunur. Ortak özellik: nükleik
KALITSAL MOLEKÜLÜN BİÇİMİ ve ORGANİZASYONU PROF. DR. SERKAN YILMAZ Değişik canlı gruplarında kalıtsal molekülün çeşidi, sayısı, biçimi ve organizasyonu bakımından farklılıklar bulunur. Ortak özellik: nükleik
ayxmaz/biyoloji Adı: 1.Aşağıda verilen atomların bağ yapma sayılarını (H) ekleyerek gösterin. C N O H
 ekleyerek gösterin. C N O H") Adı: 1.Aşağıda verilen atomların bağ yapma sayılarını (H) ekleyerek gösterin. C N O H 2.Radyoaktif izotoplar biyologları için önemlidir? Aşağıda radyoakif maddelerin kullanıldığı alanlar sıralanmıştır.bunlarla
Adı: 1.Aşağıda verilen atomların bağ yapma sayılarını (H) ekleyerek gösterin. C N O H 2.Radyoaktif izotoplar biyologları için önemlidir? Aşağıda radyoakif maddelerin kullanıldığı alanlar sıralanmıştır.bunlarla
Akıllı Defter. 9.Sınıf Biyoloji. vitaminler,hormonlar,nükleik asitler. sembole tıklayınca etkinlik açılır. sembole tıklayınca ppt sunumu açılır
 9.Sınıf Biyoloji 1 Akıllı Defter vitaminler,hormonlar,nükleik asitler sembole tıklayınca etkinlik açılır sembole tıklayınca ppt sunumu açılır sembole tıklayınca video açılır 1 VİTAMİNLER ***Vitaminler:
9.Sınıf Biyoloji 1 Akıllı Defter vitaminler,hormonlar,nükleik asitler sembole tıklayınca etkinlik açılır sembole tıklayınca ppt sunumu açılır sembole tıklayınca video açılır 1 VİTAMİNLER ***Vitaminler:
Şekil Su ürünleri yetiştiriciliği yapılan havuzlarda fosfor döngüsü (Boyd and Tucker 1998)
") - Fosfor Doğal sularda fosfor, inorganik ve organik fosfatlar (PO 4 ) halinde bulunur. Fosfor, canlı protoplazmanın yaklaşık % 2 sini oluşturduğundan yetersizliğinde, özellikle fotosentezle üretim yapan
- Fosfor Doğal sularda fosfor, inorganik ve organik fosfatlar (PO 4 ) halinde bulunur. Fosfor, canlı protoplazmanın yaklaşık % 2 sini oluşturduğundan yetersizliğinde, özellikle fotosentezle üretim yapan
YGS YE HAZIRLIK DENEMESi #6
 YGS YE HAZIRLIK DENEMESi #6 1) Canlılar soylarının devam ettirebilmek için üreyerek yeni bireyler meydana getirir. Bu üreme olaylarıyla ilgili olarak; I. Bakterinin ikiye bölünerek kendine benzer yeni
YGS YE HAZIRLIK DENEMESi #6 1) Canlılar soylarının devam ettirebilmek için üreyerek yeni bireyler meydana getirir. Bu üreme olaylarıyla ilgili olarak; I. Bakterinin ikiye bölünerek kendine benzer yeni
Ayxmaz/biyoloji. Azot döngüsü. Azot kaynakları 1. Atmosfer 2. Su 3. Kara 4. Canlılar. Azot döngüsü
 Azot döngüsü Azot kaynakları 1. Atmosfer 2. Su 3. Kara 4. Canlılar Azot döngüsü 1. Azot bitkiler tarafından organik moleküllerin (A.asit,organik baz vb.)yapısına katılır. 2. Bitkiler azotu sadece NO3-
Azot döngüsü Azot kaynakları 1. Atmosfer 2. Su 3. Kara 4. Canlılar Azot döngüsü 1. Azot bitkiler tarafından organik moleküllerin (A.asit,organik baz vb.)yapısına katılır. 2. Bitkiler azotu sadece NO3-
1. KİMYASAL ANALİZLER
 1. KİMYASAL ANALİZLER HPLC VE LC-MS/MS CİHAZLARI İLE YAPILAN ANALİZLER SORBAT TAYİNİ BENZOAT TAYİNİ KAFEİN TAYİNİ HMF TAYİNİ SUDAN TÜREVLERİ TAYİNİ VANİLİN TAYİNİ GLUKOZ, FRUKTOZ VE SUKROZ TAYİNİ SAPONİN
1. KİMYASAL ANALİZLER HPLC VE LC-MS/MS CİHAZLARI İLE YAPILAN ANALİZLER SORBAT TAYİNİ BENZOAT TAYİNİ KAFEİN TAYİNİ HMF TAYİNİ SUDAN TÜREVLERİ TAYİNİ VANİLİN TAYİNİ GLUKOZ, FRUKTOZ VE SUKROZ TAYİNİ SAPONİN
Canlının yapısında bulunan organik molekül grupları; o Karbonhidratlar o Yağlar o Proteinler o Enzimler o Vitaminler o Nükleik asitler ve o ATP
 Tamamı karbon ( C ) elementi taşıyan moleküllerden oluşan bir gruptur. Doğal organik bileşikler canlı vücudunda sentezlenir. Ancak günümüzde birçok organik bileşik ( vitamin, hormon, antibiyotik vb. )
Tamamı karbon ( C ) elementi taşıyan moleküllerden oluşan bir gruptur. Doğal organik bileşikler canlı vücudunda sentezlenir. Ancak günümüzde birçok organik bileşik ( vitamin, hormon, antibiyotik vb. )
Hücre. 1 µm = 0,001 mm (1000 µm = 1 mm)!
!") HÜCRE FİZYOLOJİSİ Hücre Hücre: Tüm canlıların en küçük yapısal ve fonksiyonel ünitesi İnsan vücudunda trilyonlarca hücre bulunur Fare, insan veya filin hücreleri yaklaşık aynı büyüklükte Vücudun büyüklüğü
HÜCRE FİZYOLOJİSİ Hücre Hücre: Tüm canlıların en küçük yapısal ve fonksiyonel ünitesi İnsan vücudunda trilyonlarca hücre bulunur Fare, insan veya filin hücreleri yaklaşık aynı büyüklükte Vücudun büyüklüğü
YGS ANAHTAR SORULAR #2
 YGS ANAHTAR SORULAR #2 1) Bir hayvan hücresinde laktoz yapımı ile ilgili olarak, sitoplazmadaki madde miktarının değişimlerini gösteren grafik aşağıdakilerden hangisi olamaz? A) Glikoz B) Su miktarı 2)
YGS ANAHTAR SORULAR #2 1) Bir hayvan hücresinde laktoz yapımı ile ilgili olarak, sitoplazmadaki madde miktarının değişimlerini gösteren grafik aşağıdakilerden hangisi olamaz? A) Glikoz B) Su miktarı 2)
BİLEŞİKLER VE FORMÜLLERİ
 BİLEŞİKLER VE FORMÜLLERİ Bileşikler : Günümüzde bilinen 117 element olmasına rağmen (92 tanesi doğada bulunur). Bu elementler farklı sayıda ve şekilde birleşerek ve etkileşerek farklı kimyasal özelliklere
BİLEŞİKLER VE FORMÜLLERİ Bileşikler : Günümüzde bilinen 117 element olmasına rağmen (92 tanesi doğada bulunur). Bu elementler farklı sayıda ve şekilde birleşerek ve etkileşerek farklı kimyasal özelliklere
FİTOPLANKTONİK ORGANİZMALARIN GENEL ÖZELLİKLERİ
 MAKALE FİTOPLANKTONİK ORGANİZMALARIN GENEL ÖZELLİKLERİ Atilla HAŞİMOĞLU, SUMAE Su kolonu içerisinde asılı olarak bulunan partiküllerden canlı organizmaları içerenlere plankton, cansız olanlara genel olarak
MAKALE FİTOPLANKTONİK ORGANİZMALARIN GENEL ÖZELLİKLERİ Atilla HAŞİMOĞLU, SUMAE Su kolonu içerisinde asılı olarak bulunan partiküllerden canlı organizmaları içerenlere plankton, cansız olanlara genel olarak
Öğr. Gör. Dr. İlker BÜYÜK (Botanik, 10. Hafta): Fotosentez FOTOSENTEZ
: Fotosentez FOTOSENTEZ") FOTOSENTEZ Elektron Koparılması ve Floresans Enerjisi Elektronlar negatif (e - ) ve protonlar pozitif (p + ) yüklüdür. Bu nedenle protonlar elektronları çekerler. Elektronlar ise, belli bir enerjiye sahiptir
FOTOSENTEZ Elektron Koparılması ve Floresans Enerjisi Elektronlar negatif (e - ) ve protonlar pozitif (p + ) yüklüdür. Bu nedenle protonlar elektronları çekerler. Elektronlar ise, belli bir enerjiye sahiptir
CANLILARIN TEMEL BİLEŞENLERİ
 1 CANLILARIN TEMEL BİLEŞENLERİ Canlıların temel bileşenleri; inorganik ve organik bileşikler olmak üzere ikiye ayrılır. **İnorganik bileşikler: Canlılar tarafından sentezlenemezler. Dışarıdan hazır olarak
1 CANLILARIN TEMEL BİLEŞENLERİ Canlıların temel bileşenleri; inorganik ve organik bileşikler olmak üzere ikiye ayrılır. **İnorganik bileşikler: Canlılar tarafından sentezlenemezler. Dışarıdan hazır olarak
YGS YE HAZIRLIK DENEMESi #22
 YGS YE HAZIRLIK DENEMESi #22 1) Zigottan başlayıp yeni bir bireyin meydana gelmesiyle sonlanan olayların hepsine birden gelişme denir. Embriyonun gelişimi sırasında, I. Morula II. Gastrula III. Blastula
YGS YE HAZIRLIK DENEMESi #22 1) Zigottan başlayıp yeni bir bireyin meydana gelmesiyle sonlanan olayların hepsine birden gelişme denir. Embriyonun gelişimi sırasında, I. Morula II. Gastrula III. Blastula
-Kloroplast ve mitokondri bulunmaz fakat bu organellerde bulunan aynı bulunur.
 BAKTERİLER GENEL ÖZELLİKLERİ: -Prokaryot hücre yapılı, tek hücreli canlılardır. -Halkasal DNA ya sahiptirler. Bazı bakterilerde plazmit bulunur. Plazmit: Küçük ve halka şeklinde DNA parçacıklarıdır. Bakterilerin
BAKTERİLER GENEL ÖZELLİKLERİ: -Prokaryot hücre yapılı, tek hücreli canlılardır. -Halkasal DNA ya sahiptirler. Bazı bakterilerde plazmit bulunur. Plazmit: Küçük ve halka şeklinde DNA parçacıklarıdır. Bakterilerin
8 HAFTA Mikrobiyal Beslenme
 Copyright McGraw-Hill companies, Inc. Permission required for reproduction or display. 8 HAFTA Mikrobiyal Beslenme 1 Copyright McGraw-Hill companies, Inc. Permission required for reproduction or display.
Copyright McGraw-Hill companies, Inc. Permission required for reproduction or display. 8 HAFTA Mikrobiyal Beslenme 1 Copyright McGraw-Hill companies, Inc. Permission required for reproduction or display.
Genel Mikrobiyoloji. Buders notunun hazırlanmasında aşağıda belirtilen kaynaktan bire bir yararlanılmıştır.
 Genel Mikrobiyoloji Buders notunun hazırlanmasında aşağıda belirtilen kaynaktan bire bir yararlanılmıştır. KAYNAK: Brock Mikroorganizmaların Biyolojisi, (Ed: Madigan, M. T., Martinko, J. M.), Çeviri Editörü:
Genel Mikrobiyoloji Buders notunun hazırlanmasında aşağıda belirtilen kaynaktan bire bir yararlanılmıştır. KAYNAK: Brock Mikroorganizmaların Biyolojisi, (Ed: Madigan, M. T., Martinko, J. M.), Çeviri Editörü:
OKSİJENLİ SOLUNUM
 1 ----------------------- OKSİJENLİ SOLUNUM ----------------------- **Oksijenli solunum (aerobik): Besinlerin, oksijen yardımıyla parçalanarak, ATP sentezlenmesine oksijenli solunum denir. Enzim C 6 H
1 ----------------------- OKSİJENLİ SOLUNUM ----------------------- **Oksijenli solunum (aerobik): Besinlerin, oksijen yardımıyla parçalanarak, ATP sentezlenmesine oksijenli solunum denir. Enzim C 6 H
MOLEKÜLER EKOTOKSİKOLOJİ LABORATUARI
 MOLEKÜLER EKOTOKSİKOLOJİ LABORATUARI Laboratuvar Sorumlusu: Prof.Dr. Mustafa CANLI İletişim : Tel: 2559, Eposta: mcanli@cu.edu.tr Temel Araştırma Alanımız: Ağır metallerin sucul organizmalara olan toksik
MOLEKÜLER EKOTOKSİKOLOJİ LABORATUARI Laboratuvar Sorumlusu: Prof.Dr. Mustafa CANLI İletişim : Tel: 2559, Eposta: mcanli@cu.edu.tr Temel Araştırma Alanımız: Ağır metallerin sucul organizmalara olan toksik
2)Subatomik parçacıklardan oluşan radyasyon. α, β ışınları
Subatomik parçacıklardan oluşan radyasyon. α, β ışınları") B) RADYASYON UYGULAMALARI Radyasyon = enerji yayılması 1)Elektromanyetik radyasyon. UV, X ve γ ışınları 2)Subatomik parçacıklardan oluşan radyasyon. α, β ışınları İyonizan ışınların canlı hücreler üzerine
B) RADYASYON UYGULAMALARI Radyasyon = enerji yayılması 1)Elektromanyetik radyasyon. UV, X ve γ ışınları 2)Subatomik parçacıklardan oluşan radyasyon. α, β ışınları İyonizan ışınların canlı hücreler üzerine
18.Eyl Rektörlük Programı Eğitim Köyü Pazartesi Rektörlük Programı Eğitim Köyü Rektörlük Programı Eğitim Köyü
 18.Eyl.17 09.00-09.50 Rektörlük Programı Eğitim Köyü Pazartesi 10.00-10.50 Rektörlük Programı Eğitim Köyü 11.00-11.50 Rektörlük Programı Eğitim Köyü 13.00-13.50 Rektörlük Programı Eğitim Köyü 14.00-14.50
18.Eyl.17 09.00-09.50 Rektörlük Programı Eğitim Köyü Pazartesi 10.00-10.50 Rektörlük Programı Eğitim Köyü 11.00-11.50 Rektörlük Programı Eğitim Köyü 13.00-13.50 Rektörlük Programı Eğitim Köyü 14.00-14.50
İÇİNDEKİLER BÖLÜM 1: MİKROBİYOLOJİYE GİRİŞ...1 BÖLÜM 2: MİKROORGANİZMALARIN MORFOLOJİLERİ.13 BÖLÜM 3: MİKROORGANİZMALARIN HÜCRE YAPILARI...
 İÇİNDEKİLER BÖLÜM 1: MİKROBİYOLOJİYE GİRİŞ...1 1.1. Tanım ve Kapsam...1 1.2. Mikrobiyoloji Biliminin Gelişmesi...2 1.3. Mikroorganizmaların Hayatımızdaki Önemi...5 1.3.1. Mikroorganizmaların Yararları...5
İÇİNDEKİLER BÖLÜM 1: MİKROBİYOLOJİYE GİRİŞ...1 1.1. Tanım ve Kapsam...1 1.2. Mikrobiyoloji Biliminin Gelişmesi...2 1.3. Mikroorganizmaların Hayatımızdaki Önemi...5 1.3.1. Mikroorganizmaların Yararları...5
(ZORUNLU) MOLEKÜLER İMMÜNOLOJİ I (TBG 607 TEORİK 3, 3 KREDİ)
 MOLEKÜLER İMMÜNOLOJİ I (TBG 607 TEORİK 3, 3 KREDİ)") T. C. İSTANBUL BİLİM ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ VE GENETİK ANABİLİM DALI YÜKSEK LİSANS PROGRAMI 2015-2016 EĞİTİM-ÖĞRETİM YILI DERS İÇERİKLERİ I. YARIYIL (ZORUNLU) MOLEKÜLER
T. C. İSTANBUL BİLİM ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ VE GENETİK ANABİLİM DALI YÜKSEK LİSANS PROGRAMI 2015-2016 EĞİTİM-ÖĞRETİM YILI DERS İÇERİKLERİ I. YARIYIL (ZORUNLU) MOLEKÜLER
DENİZ BALIKLARI YETİŞTİRİCİLİĞİNDE SU KALİTESİ
 DENİZ BALIKLARI YETİŞTİRİCİLİĞİNDE SU KALİTESİ Su ürünleri yetiştiriciliği açısından önemli su kalite özellikleri ve bu özelliklere ilişkin sınır (standart) değerler uzun yıllar süren araştırma ve deneyimler
DENİZ BALIKLARI YETİŞTİRİCİLİĞİNDE SU KALİTESİ Su ürünleri yetiştiriciliği açısından önemli su kalite özellikleri ve bu özelliklere ilişkin sınır (standart) değerler uzun yıllar süren araştırma ve deneyimler
Sitoplazmik membran periferal integral
 İÇ YAPILAR Sitoplazmik membran Hücre duvarının altında, ince ve bakterilerde genellikle aynı yapıda İki katmandan oluşur Periplasmik boşluk ve sitoplazmaya bakan yüzeyde, protein ve fosfolipid İç bölgede,
İÇ YAPILAR Sitoplazmik membran Hücre duvarının altında, ince ve bakterilerde genellikle aynı yapıda İki katmandan oluşur Periplasmik boşluk ve sitoplazmaya bakan yüzeyde, protein ve fosfolipid İç bölgede,
PROKARYOT VE ÖKARYOT HÜCRELER
 PROKARYOT VE ÖKARYOT HÜCRELER HÜCRE Hücre ya da göze, bir canlının yapısal ve işlevsel özellikleri gösterebilen en küçük birimidir. Hücre, (İng. Cell); Latince küçük odacık anlamına gelen "cellula" kelimesinden
PROKARYOT VE ÖKARYOT HÜCRELER HÜCRE Hücre ya da göze, bir canlının yapısal ve işlevsel özellikleri gösterebilen en küçük birimidir. Hücre, (İng. Cell); Latince küçük odacık anlamına gelen "cellula" kelimesinden
BİYOKİMYAYA GİRİŞ: ATOM, MOLEKÜL, ORGANİK BİLEŞİKLER
 BİYOKİMYAYA GİRİŞ: ATOM, MOLEKÜL, ORGANİK BİLEŞİKLER Biyokimyanın tanımı yaşamın temel kimyası ile ilgilenen bilim dalı (Bios, Yunancada yaşam demektir.) canlı sistemin yapısını ve fonksiyonlarını kimyasal
BİYOKİMYAYA GİRİŞ: ATOM, MOLEKÜL, ORGANİK BİLEŞİKLER Biyokimyanın tanımı yaşamın temel kimyası ile ilgilenen bilim dalı (Bios, Yunancada yaşam demektir.) canlı sistemin yapısını ve fonksiyonlarını kimyasal
Mikrobiyal Gelişim. Jenerasyon süresi. Bakterilerde üreme eğrisi. Örneğin; (optimum koşullar altında) 10/5/2015
 10/5/2015") Mikrobiyal Gelişim Tek hücreli organizmalarda sayı artışı Bakterilerde en çok görülen üreme şekli ikiye bölünmedir (mikroorganizma sayısı) Çok hücreli organizmalarda kütle artışı Genelde funguslarda görülen
Mikrobiyal Gelişim Tek hücreli organizmalarda sayı artışı Bakterilerde en çok görülen üreme şekli ikiye bölünmedir (mikroorganizma sayısı) Çok hücreli organizmalarda kütle artışı Genelde funguslarda görülen
11. SINIF KONU ANLATIMI 4 FOTOSENTEZ - 2 FOTOSENTEZDE GÖREV ALAN YAPILAR
 11. SINIF KONU ANLATIMI 4 FOTOSENTEZ - 2 FOTOSENTEZDE GÖREV ALAN YAPILAR Ökaryotik canlılarda klorofil pigmentini taşıyan plastidtir. Fotosentezle görevlidir. Kloroplastta dış ve iç olmak üzere iki tane
11. SINIF KONU ANLATIMI 4 FOTOSENTEZ - 2 FOTOSENTEZDE GÖREV ALAN YAPILAR Ökaryotik canlılarda klorofil pigmentini taşıyan plastidtir. Fotosentezle görevlidir. Kloroplastta dış ve iç olmak üzere iki tane
Metschnikowia pulcherrima Türü Mayaların İzolasyonu ve Pulcherrimin in Antimikrobiyal Aktivitelerinin Araştırılması. Prof. Dr.
 Metschnikowia pulcherrima Türü Mayaların İzolasyonu ve Pulcherrimin in Antimikrobiyal Aktivitelerinin Araştırılması Prof. Dr. Sezai Türkel Uludağ Üniversitesi, Fen-Edebiyat Fakültesi, Biyoloji Bölümü,
Metschnikowia pulcherrima Türü Mayaların İzolasyonu ve Pulcherrimin in Antimikrobiyal Aktivitelerinin Araştırılması Prof. Dr. Sezai Türkel Uludağ Üniversitesi, Fen-Edebiyat Fakültesi, Biyoloji Bölümü,
13 HÜCRESEL SOLUNUM LAKTİK ASİT FERMANTASYONU
 13 HÜCRESEL SOLUNUM LAKTİK ASİT FERMANTASYONU Laktik Asit Fermantasyonu Glikozdan oksijen yokluğunda laktik asit üretilmesine LAKTİK ASİT FERMANTASYONU denir. Bütün canlılarda sitoplazmada gerçekleşir.
13 HÜCRESEL SOLUNUM LAKTİK ASİT FERMANTASYONU Laktik Asit Fermantasyonu Glikozdan oksijen yokluğunda laktik asit üretilmesine LAKTİK ASİT FERMANTASYONU denir. Bütün canlılarda sitoplazmada gerçekleşir.
Günümüzde bilinen 117 element olmasına rağmen (92 tanesi doğada bulunur) bu elementler farklı sayıda ve şekilde birleşerek ve etkileşerek farklı
 bu elementler farklı sayıda ve şekilde birleşerek ve etkileşerek farklı") Günümüzde bilinen 117 element olmasına rağmen (92 tanesi doğada bulunur) bu elementler farklı sayıda ve şekilde birleşerek ve etkileşerek farklı kimyasal özelliklere sahip milyonlarca yani madde yani bileşik
Günümüzde bilinen 117 element olmasına rağmen (92 tanesi doğada bulunur) bu elementler farklı sayıda ve şekilde birleşerek ve etkileşerek farklı kimyasal özelliklere sahip milyonlarca yani madde yani bileşik
SALGIN ARAŞTIRMASINDA KULLANILAN TİPLENDİRME YÖNTEMLERİ
 SALGIN ARAŞTIRMASINDA KULLANILAN TİPLENDİRME YÖNTEMLERİ Prof.Dr. Meltem Yalınay Çırak Gazi Üniversitesi Tıp Fakültesi Mikrobiyoloji ve Klinik Mikrobiyoloji A.D. fenotipik yöntemler genotipik yöntemler
SALGIN ARAŞTIRMASINDA KULLANILAN TİPLENDİRME YÖNTEMLERİ Prof.Dr. Meltem Yalınay Çırak Gazi Üniversitesi Tıp Fakültesi Mikrobiyoloji ve Klinik Mikrobiyoloji A.D. fenotipik yöntemler genotipik yöntemler
7. BÖLÜM MİKROBİYAL GELİŞİM
 7. BÖLÜM MİKROBİYAL GELİŞİM 1 Gelişim Tek hücreli organizmalarda sayı artışı Bakterilerde en çok görülen üreme şekli ikiye bölünmedir (mikroorganizma sayısı) Çok hücreli organizmalarda kütle artışı Genelde
7. BÖLÜM MİKROBİYAL GELİŞİM 1 Gelişim Tek hücreli organizmalarda sayı artışı Bakterilerde en çok görülen üreme şekli ikiye bölünmedir (mikroorganizma sayısı) Çok hücreli organizmalarda kütle artışı Genelde
GÖRÜNÜR IŞIĞIN HAVUZ SULARININ DEZENFEKSİYONUNDA ALTERNATİF BİR YÖNTEM OLARAK KULLANILMASI
 GÖRÜNÜR IŞIĞIN HAVUZ SULARININ DEZENFEKSİYONUNDA ALTERNATİF BİR YÖNTEM OLARAK KULLANILMASI Hazırlayan Öğrenciler Dila Berfin UÇAN 7-F Ekin Ladin TÜRKMEN 7-F Danışman Öğretmen Melike TURAN İZMİR, 2014 İÇİNDEKİLER
GÖRÜNÜR IŞIĞIN HAVUZ SULARININ DEZENFEKSİYONUNDA ALTERNATİF BİR YÖNTEM OLARAK KULLANILMASI Hazırlayan Öğrenciler Dila Berfin UÇAN 7-F Ekin Ladin TÜRKMEN 7-F Danışman Öğretmen Melike TURAN İZMİR, 2014 İÇİNDEKİLER
Hücrede Genetik Bilgi Akışı
 Hücrede Genetik Bilgi Akışı 1) Genomun korunması DNA nın tam olarak kopyalanması ve hücre bölünmesiyle yeni kuşak hücrelere aktarılması 2) Genetik bilginin çevrimi Hücre içerisinde bilginin DNA dan RNA
Hücrede Genetik Bilgi Akışı 1) Genomun korunması DNA nın tam olarak kopyalanması ve hücre bölünmesiyle yeni kuşak hücrelere aktarılması 2) Genetik bilginin çevrimi Hücre içerisinde bilginin DNA dan RNA
TRANSKRİPSİYON AŞAMASINDA KROMATİN YAPININ DÜZENLENMESİ
 İ.Ü Fen Bilimleri Enstitüsü Moleküler Biyoloji ve Genetik TRANSKRİPSİYON AŞAMASINDA KROMATİN YAPININ DÜZENLENMESİ Merve YILMAZER 2601120219 İÇERİK Kromatin ve nükleozom yapısı Transkripsiyon aşamasında
İ.Ü Fen Bilimleri Enstitüsü Moleküler Biyoloji ve Genetik TRANSKRİPSİYON AŞAMASINDA KROMATİN YAPININ DÜZENLENMESİ Merve YILMAZER 2601120219 İÇERİK Kromatin ve nükleozom yapısı Transkripsiyon aşamasında
SU ÜRÜNLERİ SAĞLIĞI BÖLÜM BAŞKANLIĞI
 SU ÜRÜNLERİ SAĞLIĞI BÖLÜM BAŞKANLIĞI Hacı SAVAŞ-SÜMAE, Su Ürünleri Sağlığı Bölüm Başkanı Su Ürünleri Sağlığı Bölüm Başkanlığı enstitümüz bünyesinde faaliyet gösteren bölümlerden birisidir. 2000 yılı başından
SU ÜRÜNLERİ SAĞLIĞI BÖLÜM BAŞKANLIĞI Hacı SAVAŞ-SÜMAE, Su Ürünleri Sağlığı Bölüm Başkanı Su Ürünleri Sağlığı Bölüm Başkanlığı enstitümüz bünyesinde faaliyet gösteren bölümlerden birisidir. 2000 yılı başından
Referans:e-TUS İpucu Serisi Biyokimya Ders Notları Sayfa:368
 21. Aşağıdakilerden hangisinin fizyolojik ph'de tamponlama etkisi vardır? A) CH3COC- / CH3COOH (pka = 4.76) B) HPO24- / H2PO-4 (pka = 6.86) C) NH3/NH+4(pKa =9.25) D) H2PO-4 / H3PO4 (pka =2.14) E) PO34-/
21. Aşağıdakilerden hangisinin fizyolojik ph'de tamponlama etkisi vardır? A) CH3COC- / CH3COOH (pka = 4.76) B) HPO24- / H2PO-4 (pka = 6.86) C) NH3/NH+4(pKa =9.25) D) H2PO-4 / H3PO4 (pka =2.14) E) PO34-/
Yrd.Doç.Dr. Yosun MATER
 * Yrd.Doç.Dr.Yosun MATER Yrd.Doç.Dr. Yosun MATER *Bitki nüklear, mitokondriyal ve kloroplast DNA'ları *Burada yer alan bugünkü bilgilerimizin çoğu, moleküler evrim mekanizması ve oranları kullanılarak
* Yrd.Doç.Dr.Yosun MATER Yrd.Doç.Dr. Yosun MATER *Bitki nüklear, mitokondriyal ve kloroplast DNA'ları *Burada yer alan bugünkü bilgilerimizin çoğu, moleküler evrim mekanizması ve oranları kullanılarak
Proteinler. Fonksiyonlarına göre proteinler. Fonksiyonlarına göre proteinler
 Proteinler Canlılarda miktar olarak en çok bulunan biyomoleküllerdir. Amino asit birimlerinden oluşurlar Yapısal ve işlevsel olabilirler Genlerle aktarılan kalıtsal bilginin ortaya çıktığı moleküllerdir.
Proteinler Canlılarda miktar olarak en çok bulunan biyomoleküllerdir. Amino asit birimlerinden oluşurlar Yapısal ve işlevsel olabilirler Genlerle aktarılan kalıtsal bilginin ortaya çıktığı moleküllerdir.
Laboratuvar Tekniği. Adnan Menderes Üniversitesi Tarımsal Biyoteknoloji Bölümü TBY 118 Muavviz Ayvaz (Yrd. Doç. Dr.) 5. Hafta (14.03.
 5. Hafta (14.03.") Laboratuvar Tekniği Adnan Menderes Üniversitesi Tarımsal Biyoteknoloji TBY 118 Muavviz Ayvaz (Yrd. Doç. Dr.) 5. Hafta (14.03.2014) 1 5. Haftanın Ders İçeriği DNA ekstraksiyonu DNA ekstraksiyonunun amacı
Laboratuvar Tekniği Adnan Menderes Üniversitesi Tarımsal Biyoteknoloji TBY 118 Muavviz Ayvaz (Yrd. Doç. Dr.) 5. Hafta (14.03.2014) 1 5. Haftanın Ders İçeriği DNA ekstraksiyonu DNA ekstraksiyonunun amacı
YAZILIYA HAZIRLIK TEST SORULARI. 11. Sınıf
 YAZILIYA HAZIRLIK TEST SORULARI 11. Sınıf 1) Oksijenli solunumda, oksijen molekülleri, I. Oksidatif fosforilasyon II. Glikoliz II. Krebs Evrelerinden hangilerinde kullanılır? A) Yalnız I B) Yalnız II C)
YAZILIYA HAZIRLIK TEST SORULARI 11. Sınıf 1) Oksijenli solunumda, oksijen molekülleri, I. Oksidatif fosforilasyon II. Glikoliz II. Krebs Evrelerinden hangilerinde kullanılır? A) Yalnız I B) Yalnız II C)
Protein Ekstraksiyonu
 Protein Ekstraksiyonu Dr.Gaye Güler Tezel Hacettepe Üniversitesi Tıp Fakültesi Patoloji Anabilim Dalı Proteinler tüm canlı organizmalar için en önemli makromoleküllerden biridir. Bazıları yapısal komponentleri
Protein Ekstraksiyonu Dr.Gaye Güler Tezel Hacettepe Üniversitesi Tıp Fakültesi Patoloji Anabilim Dalı Proteinler tüm canlı organizmalar için en önemli makromoleküllerden biridir. Bazıları yapısal komponentleri
ADIM ADIM YGS-LYS 43. ADIM CANLILARIN SINIFLANDIRILMASI-3 BAKTERİLER ALEMİ
 ADIM ADIM YGS-LYS 43. ADIM CANLILARIN SINIFLANDIRILMASI-3 BAKTERİLER ALEMİ BAKTERİLER ALEMİ Prokaryot hücre yapısına sahip olan tek hücreli canlıların oluşturduğu alemdir. Mikroskobun icadından sonra keşfedilmişlerdir.
ADIM ADIM YGS-LYS 43. ADIM CANLILARIN SINIFLANDIRILMASI-3 BAKTERİLER ALEMİ BAKTERİLER ALEMİ Prokaryot hücre yapısına sahip olan tek hücreli canlıların oluşturduğu alemdir. Mikroskobun icadından sonra keşfedilmişlerdir.
ARAŞTIRMA ENSTİTÜSÜ/İSTASYONLARI MÜDÜRLÜKLERİ DÖNER SERMAYE İŞLETMELERİ 2014 YILI BİRİM FİYAT LİSTESİ. 1 ph 14,00. 2 Elektriksel İletkenlik 14,00
 ARAŞTIRMA ENSTİTÜSÜ/İSTASYONLARI MÜDÜRLÜKLERİ DÖNER SERMAYE İŞLETMELERİ 2014 YILI BİRİM FİYAT LİSTESİ Sıra No: SULAMA SUYU ANALİZLERİ: 2014 FİYATI 1 ph 14,00 2 Elektriksel İletkenlik 14,00 3 Sodyum (Na)
ARAŞTIRMA ENSTİTÜSÜ/İSTASYONLARI MÜDÜRLÜKLERİ DÖNER SERMAYE İŞLETMELERİ 2014 YILI BİRİM FİYAT LİSTESİ Sıra No: SULAMA SUYU ANALİZLERİ: 2014 FİYATI 1 ph 14,00 2 Elektriksel İletkenlik 14,00 3 Sodyum (Na)