Zir.Yük.Müh.Yıldız D LL Bilim Dalı Kodu: Sunu Tarihi: 30 / 06/ 2008
|
|
|
- Onur Karadere
- 5 yıl önce
- İzleme sayısı:
Transkript
1 I EGE ÜN VERS TES FEN B L MLER ENST TÜSÜ (DOKTORA TEZ ) EGE BÖLGES NDEK BAZI ÖNEML ÜZÜM ÇE TLER, T PLER VE KLONLARININ M KROSATEL T (SSR) MARKÖRLER YLE KARAKTER ZASYONU ÜZER NDE ARA TIRMALAR Zir.Yük.Müh.Yıldız D LL Bilim Dalı Kodu: Sunu Tarihi: 30 / 06/ 2008 Tez Danı manı: Prof. Dr. Ahmet ALTIND L Bornova ZM R
2 II
3 III Sayın Yıldız D LL tarafından Doktora tezi olarak sunulan Ege Bölgesindeki Bazı Önemli Üzüm Çe itleri, Tipleri ve Klonlarının Mikrosatelit (SSR) Markörleriyle Karakterizasyonu Üzerinde Ara tırmalar ba lıklı bu çalı ma, E.Ü. Fen Bilimleri Enstitüsü E itim ve Ö retim Yönetmeli i ile E.Ü. Fen Bilimleri Enstitüsü E itim ve Ö retim Yönergesi nin ilgili hükümleri uyarınca tarafımızdan de erlendirilerek savunmaya de er bulunmu ve 30/06/2008 tarihinde yapılan tez savunma sınavında aday oybirli i ile ba arılı bulunmu tur. Jüri Üyeleri: mza Jüri Ba kanı: Prof. Dr. Ahmet ALTIND L Üye: Doç. Dr. Sacide PEHL VAN Üye: Doç. Dr. Ali ERGÜL Üye: Doç. Dr. Hülya LB Üye: Yrd. Doç. Dr. Harun ÇOBAN
4 IV
5 V ÖZET EGE BÖLGES NDEK BAZI ÖNEML ÜZÜM ÇE TLER, T PLER VE KLONLARININ M KROSATEL T (SSR) MARKÖRLER YLE KARAKTER ZASYONU ÜZER NDE ARA TIRMALAR D LL, Yıldız Doktora Tezi, Bahçe Bitkileri Anabilim Dalı Tez Yöneticisi: Prof. Dr. Ahmet ALTIND L Haziran 2008, 96 Sayfa Bu ara tırmada, Manisa Ba cılık Ara tırma Enstitüsü nce klon seleksiyonu sonucu seçilmi Sultani Çekirdeksiz üzüm çe idine ait 5 tip, Pembe Gemre, Osmanca ve pek üzüm çe itlerine ait 9 klon ve Ege Bölgesi için önemi olan 15 yerel çe it ile 2 referans çe it olmak üzere toplam 31 üzüm çe idinin (Vitis vinifera L.) SSR markörleri ile DNA düzeyinde allel profilleriyle genetik tanımlaması yapılarak aralarındaki genetik benzerlikler ve farklılıklar belirlenmi tir. Lokuslarda gözlenen allel sayısı 6 ile 15 arasında (VVS3, VrZAG62) de i mi tir. Beklenen ve gözlenen heterozigotluk oranları sırasıyla ortalama ve olarak bulunmu tur. Kümeleme analizi için UPGMA (Unweighted Pair- Group Method Using Arithmetic Means) yöntemi uygulanmı tır. Dendogramda 2 ana grup ortaya çıkmı tır. Genotiplerin büyük kısmını içeren ilk grup içinde çok sayıda alt grup gözlenmi tir. kinci grupta ise Osmanca çe idinin 26, 39 ve 40 no lu klonları ile birlikte yerli araplık çe itlerimizden Foça Karası (09), Foça Karası (17), Foça Karası (M) çe itleri birlikte yer almı tır. Genotipler arasında Aydın, Çanakkale ve
6 VI zmir (Mordo an) illerinden alınan Foça Karası çe itleri ve Manisa Ba cılık Ara tırma Enstitüsü nde farklı parsellerde yer alan Dumanlı Gemre çe itlerinde de homonim duruma rastlanmı tır. Sultani Çekirdeksiz çe idinin kurutmalık özelli e sahip tiplerinden T3, T15, Y3 ile sofralık özelliklere sahip S4 ve S6 tiplerinin genetik olarak birbirlerinden farklı oldu u ortaya konmu tur. Anahtar Kelimeler : Vitis vinifera L., asma, mikrosatelit, genetik tanımlama
7 VII ABSTRACT RESEARCHES ON THE CHARACTERIZATION OF SOME IMPORTANT GRAPE VARIETIES, TYPES AND CLONES IN THE AEGEAN REGION BY USING MICROSATELLITE (SSR) MARKERS D LL, Yıldız Ph.D. Thesis, Department of Horticulture Supervisor: Prof. Dr. Ahmet ALTIND L June 2008, 96 pages In this study, a total of 31 grape cultivars (Vitis vinifera L.) consisting of 5 types Sultani Çekirdeksiz, 9 clones of Pembe Gemre, Osmanca and Ipek cultivars selected by Manisa Viticulture Research Institute and 15 local grape varieties plus 2 reference cultivars of economic importance in the Aegean Region were genetically characterized and genetic relationship among them were investigated at DNA level with allele profiles by using co-dominant markers of SSR (Simple Sequence Repeats). Number of alleles per locus ranged from 6 to 15 (VVS3, VrZAG62). Expected and observed heterozygosity values were and , respectively. Cluster analysis for the genotypes was conducted by using UPGMA (Unweighted Pair-Group Method Using Arithmetic Means) method. Two main groups appeared in the dendogram. Many sub-groups were observed in Group 1 that includes most of the genotypes. On the other hand, Clones 26, 39 and 40 of Osmanca, and 09, 17 and M varieties of Foça Karası, one of the local
8 VIII wine varieties in the Aegean Region, appeared in Group 2. One case of homonymy was revealed for the genotypes of Foça Karası collected from Aydın, Çanakkale, zmir (Mordo an), and Dumanlı Gemre variety which is located in the different research plots at Manisa Viticulture Research Institute. Genetic differences were determined between T3, T15 and Y3 types, which have dried grape characteristics, and S4 and S6 types, which have table grape characteristics of Sultani Çekirdeksiz variety. Key words: Vitis vinifera L., grape, microsatellite, genetic identification
9 IX TE EKKÜR Bu konuda bana çalı ma olana ı veren ve tezin yürütülmesinin her a amasında de erli katkı ve görü lerini esirgemeyen danı man hocam Sayın Prof. Dr. Ahmet ALTIND L ye, tez izleme komite üyeleri de erli hocalarım Prof. Dr. Gökhan SÖYLEMEZO LU (A.Ü.Z.F. Bahçe Bitkileri Bölümü) ve Doç.Dr Sacide PEHL VAN a (Gaziantep Ü.Tıp Fakültesi Tıbbi Biyoloji A.B.D.) da tezin çe itli a amalarında yapmı oldukları de erli katkılarından dolayı te ekkürü bir borç bilirim. Tezimdeki tekni i ö reten Sayın Hocam Doç. Dr. Ali ERGÜL e (A.Ü. Biyoteknoloji Enstitüsü), yardıma ihtiyaç duydu umda hep yanımda olan arkada ım Dr. Burçak Ç ye özel te ekkürlerimi sunarım. Tez çalı mam sırasında katkılarından dolayı Ege Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümünden Prof. Dr. brahim KISMALI, Prof. Dr. Uygun AKSOY, Prof. Dr. Serra HEPAKSOY, Doç.Dr. Hülya LB, Dr. Fatma AYKUT (E.Ü. Tarla Bitkileri Bölümü), Ar.Gör. Mert SUDA IDAN ( zmir Yüksek Teknoloji Enstitüsü), Ar. Gör. Melike BAKIR (A.Ü. Biyoteknoloji Enstitüsü) ve Eda KARAA AÇ a te ekkür ederim. Ara tırmanın yürütülmesi için gerekli finansal deste i sa layan Tarım ve Köyi leri Bakanlı ı Tarımsal Ara tırmalar Genel Müdürlü ü ne, Manisa Ba cılık Ara tırma Enstitüsü nden Dr. Adem YA CI, Dr. Selçuk KARABAT ve Dr. Sumru KADER a te ekkür ederim. Bu çalı mam ve hayatım boyunca her türlü maddi ve manevi desteklerini esirgemeyen, canım aileme en içten sevgi ve te ekkürlerimi sunuyorum.
10 X
11 XI Ç NDEK LER Sayfa ÖZET V ABSTRACT. VII TE EKKÜR. IX Ç ZELGELER D Z N.. XIII EK LLER D Z N XIV KISALTMALAR D Z N... XV 1. G R 1 2. ÖNCEK ÇALI MALAR Bitkilerin Tanımlanmasında Kullanılan Markör Tipleri Asmada Günümüze Kadar Yapılan Tanımlama Çalı maları Ampelografik Tanımlamalar zoenzim (Biyokimyasal) Tanımlamalar DNA Markörleri DNA Markörleriyle Yapılan Tanımlamalar Asmada Mikrosatelit (SSR) Markörlerin Geli tirilmesi Asmalarda Mikrosatellit Çalı maları SSR Tekni inin Klon ve Tiplerin Tanımlanmasında Kullanımı MATERYAL VE YÖNTEM Materyal Yöntem.. 41
12 XII DNA izolasyonu PCR uygulaması Çalı mada Kullanılan SSR Primerleri Agaroz Jel Elektroforezi Poliakrilamid Jel Elektroforezi ve Jel Boyama Allel De erlerinin Belirlenmesi ve Genetik Analizler ARA TIRMA BULGULARI DNA zolasyonu Allel Büyüklükleri Yazılım Programları ile Sonuçların De erlendirilmesi TARTI MA VE SONUÇ. 68 KAYNAKLAR D Z N.. 77 EKLER. 92 ÖZGEÇM. 96
13 XIII Ç ZELGELER D Z N Sayfa Çizelge 2.1. Moleküler markör tekniklerinin kar ıla tırılması... 9 Çizelge 3.1. Ara tırmada kullanılan üzüm çe itlerine ili kin kısa ampelografik özellikler Çizelge 3.2. PCR ço altma ö eleri konsantrasyon ve miktarları.. 44 Çizelge 3.3. Primerlerin PCR daki döngü ko ulları.. 44 Çizelge 3.4. Ara tırmada kullanılan primerlere ait bilgiler 45 Çizelge 4.1. Ara tırmada kullanılan üzüm çe itlerine ait DNA miktarı ve saflık dereceleri. 49 Çizelge 4.2. Çe it, tip ve klonların 15 lokustaki allel büyüklükleri. 52 Çizelge 4.3. Çalı ılan lokuslardaki allel sayıları (N), beklenen heterozigotluk (He), gözlenen heterozigotluk (Ho), tespit olasılı ı (PI) de eri ve sessiz (null) allel frekansı 57 Çizelge 4.4. Ege Bölgesine ait bazı önemli üzüm çe itleri,tipleri ve klonlarının genetik benzerlik.. 66
14 XIV EK LLER D Z N Sayfa ekil Ara tırmada Kullanılan Üzüm Çe itleri. 33 Sekil 4.1. Çalı ılan lokuslarda tespit edilen allel frekansları.. 59 ekil 4.2 Ege Bölgesi asma genotiplerine ait dendogram. 67
15 XV S MGELER VE KISALTMALAR D Z N AFLP : Amplified Fragment Length Polymorphism (Ço altılan Parça Uzunluk Farklılı ı) bp : Base pair (Baz çifti) CTAB : Hekzadesil Trimetil-Amonyum Bromür DNA : Deoksiribo Nükleik Asit dntp : Deoksi-Nüklezid Trifosfat EDTA : Etilen Diamine Tetra Asetik Asit He : Expected heterozigosity (Beklenen heterozigotluk) Ho : Observed heterozigosity (Gözlenen heterozigotluk) Kol : Koleksiyon parseli M : Mol MgCl 2 : Magnezyum Klorür µl : Mikrolitre mm : Milimol n : The number of allelles (Allel sayısı) OIV : Organisation Internationale de la Vigne et du Vin (Uluslararası Ba cılık ve arapçılık Organizasyonu) Os : Osmanca parseli PCR : Polymerase Chain Reaction (Polimeraz Zincir Reaksiyonu) PI Probability of Identity (Tespit olasılı ı) PVP : Polyvinylpyrrolidone
16 XVI r : The estimated frequency of null allele (Tahmin edilen sessiz allel frekansı) RAPD : Random Amplified Polymorphism DNA (Rastgele Ço altılmı DNA Farklılı ı) RFLP : Restriction Fragment Length Polymorphism (Kesilmi Parça Uzunlu u Farklılı ı) RNase : Ribonükleaz rpm : Dakikadaki dönü sayısı SSR : Simple Sequence Repeats (Basit Dizi Tekrarları) TBE : Tris-Borik Asit-EDTA Çözeltisi TE : Tris-EDTA Çözeltisi UPOV : The International Union for the Protection of New Varieties of Plants (Yeni Bitki Çe itlerinin Korunmasına li kin Uluslararası Birlik)
17 1 1. G R Ba cılık yeryüzünde en eski kültüre sahip tarım kollarından olup, günümüzde de önemini koruyarak kuzey ve güney yarımkürede en yaygın kültür alanlarından birini olu turmu tur. Ço u ülkede üzüm (Vitis vinifera L.) önemli bir ürün olup, en az üzüm çe idinin oldu u belirlenmi tir. Fakat gerçek sayısının çok daha az oldu u ( arasında) bilinmektedir (Alleweldt, 1988). Ba cılık; Kuzey Yarımkürede ve Güney Yarımkürede enlem dereceleri arasındaki ülkelerde yayılmı tır. Kuzey Yarımkürede, ilgili enlem dereceleri arasındaki ülkemiz, dünyanın en elveri li iklim ku a ında yer alması nedeniyle; asmanın gen merkezi ve ilk kültüre alındı ı co rafyanın merkezi konumundadır (Oraman, 1970). XX. yüzyıla kadar asmanın gen merkezi olarak Kafkasya, Hazar Denizi nin güneyi, kuzeydo u Anadolu nun da içinde oldu u Küçük Asya gösterilmi tir. Bu yüzyılda yapılan bazı ara tırmalar asmanın ilk görüldü ü zamanları, günümüzden yakla ık milyon sene öncesine kadar götürmekte ve bulundu u yerler olarak da dünyanın birçok yöresini göstermektedir (Gollmick ve ark, 1970; A ao lu ndan, 1999). Ülkemizde her bölgede yapılan kazılarda ba cılıkla ilgili tarih öncesi devirlere ait önemli eserler bulunmu tur. zmir in Bergama ilçesinde M.Ö. 3. yüzyıla ait bir kabartmada arap tanrısı Dionysos u elinde bir salkımla gösteren bir seramik eserin bulunması ve ülkemizin birçok yerinde yapılan kazılarda üzüm ve ba la ilgili benzer motiflerin elde edilmesi, ba cılı ın bu bölgede geçmi te önemli derecede geli mi
18 2 oldu unu gösteren en önemli göstergelerdir. Bugün bile Bergama da Kozak yaylasında son turfanda ve kaliteli üzümlerden Kozak Beyazı ve Kozak Siyahı üzüm çe itleri yaygın olarak yeti tirilmektedir. Ege ve Marmara Bölgesi nde ba cılı ın geli ti i yörelerde (Lapseki, Çanakkale, Bergama, Alia a, Dikili, Bozcaada, Çe me, Karaburun ve Seferihisar) basılan paralar üzerinde üzüme, arap kabına ve amfora ya yer verilmi olması, ba cılı a ve araba verilen önemi göstermektedir (Çelik, 1998). Ba cılık geçmi ten günümüze daima varolmu ve tarımsal yapı içinde önemini daima korumu tur. Ülkemiz ba cılı ının geli tirilmesi ve milli ekonomiye olan katkısının daha yüksek düzeylere ula tırılması, her eyden önce sahip oldu umuz asma gen potansiyelinin belirlenmesi, bu potansiyelin kataloglandırılarak uluslararası geçerlilikte korunabilmesi ve de erlendirilmesine yönelik olan çalı malara gereken önemin verilmesi ile mümkündür. Türkiye nin tamamının taranması tamamlanmamı olmasına ra men, Tekirda daki Milli Koleksiyon Ba ı nda 1172 adet üzüm çe idi yer almaktadır. Bu sayının fazlalı ı ülkemiz asma gen potansiyelinin ne kadar zengin oldu unu göstermektedir. Bu zenginlik gerek ıslah çalı malarında gerekse ekonomik öneme sahip yerel çe itlerin ortaya konmasında çok önemli bir kaynaktır. Üzüm çe itlerinin do ru tanımlanması; bitkilerin ismine do rulu unun de erlendirilmesi, uluslararası gen kaynakları de i iminde, yanlı isimlendirmelerin önlenmesi ve asma fidanı sertifikasyon sisteminde kritik öneme sahiptir.
19 3 Ampelografi, üzüm çe itlerinin tanımlamasında yıllardır kullanılan geleneksel bir yöntemdir. Bununla birlikte yalnız morfolojik özellikleri baz alan bu botanik esaslı metot çevresel etkiye maruz kalır ve benzer görünü lü çe itleri seçmede ba arısız olur. Ayrıca ki isel de erlendirmelere yol açtı ı gibi, hastalık ve zararlılar veya kültürel uygulamaların etkisiyle asmanın yalnızca dı görünümünde veya ölçülebilir karakterlerinde meydana gelen varyasyonların ço u kez yeni genotipler olarak adlandırılmasına neden olmaktadır (Najafi et. al., 2006; Marasalı, 1995). Asma tür ve çe itlerinin tanımlanması daha çok klasik ampelografik tanımlamalar ile yapılırken, son yıllarda DNA ya dayalı markörler; tür ve çe itlerin ayrım ve tanımlamalarında kesin sonuç vermeleri nedeni ile klasik yöntemler ile birlikte kullanılmaya ba lanmı ve bu teknikler asma genotiplerinin tanımlanmasında çok önemli ilerlemeler sa lamı tır. Biyoteknoloji alanında kaydedilen ilerlemelerle, çe itlerin kesin te hisine yardımcı olacak DNA düzeyinde moleküler markör teknikleri geli tirilmi tir. Ampelografik çalı malar; izoenzim tanımlamaları, RFLP (Restriction Fragment Length Polymorphism), RAPD (Random Amplified Polymorphic DNA), AFLP (Amplified Length Polymorphism) ve SSR (Simple Sequence Repeat) gibi teknikleri de kapsamaktadır. Bunlardan SSR markörler, polimorfizm oranlarının yüksek olu u, tekrarlanabilirli i ve kodominant karakterli olmasından dolayı tercih edilirler. Dünyada yaygın bir ekilde kullanılan SSR DNA analiz tekni inin a ırlıklı çalı ma konuları anaç, çe it, klon ve ebeveyn
20 4 tanımlamalarıdır (Oriana ve ark., 1997; Sefc ve ark., 1997; Sefc ve ark.,1998; Regner ve ark., 2000). DNA parmak izi analizi, genetik materyallerin birbiriyle benzerlik veya farklılıklarının saptanması amacını ta ır. Genetik markörlerdeki varyasyonun genlerdeki varyasyonu temsil etti i öngörüsünden hareketle parmak izi analizinde markörlerin kullanımı fikri do mu tur. Parmak izi analizinde aynı anda genomun pek çok yerine dair bilgi sa layan markörlerin kullanılması yaygındır. Sonuçta genetik materyaller arasında farklılık gösteren lokusların oranı belirlenir. Farklılı ın hangi lokusta oldu una bakılmaz. Bu tip bir analiz bitki ıslahı programına alınacak materyallerin seçiminde kullanılır ve varyasyonu yüksek olan hatların kullanımı ile ıslahçı daha geni bir de i im içinden istediklerini seçme ansına sahip olur (Brown ve Kresovich, 1996;. Özcan ve ark. dan, 2001). Ülkemizin çe it düzeyinde gen potansiyeli olmasının yanı sıra, bir çe itten elde edilen bitki toplulu u, bazı özellikler bakımından incelendi i zaman bireyler arasında varyasyonların var oldu u görülmektedir. Klon seleksiyonu yolu ile bir çe it içerisinde var olan varyasyonlardan yararlanarak çe itli özellikler bakımından üstünlük gösteren tipler seçilmektedir. Bir çe it içerisindeki populasyonlardaki farklıla manın kayna ı büyük ölçüde mutasyonlar olmaktadır. Klon seleksiyonundan amaç esas çe itten üstün de eri olan ve ekonomik de eri olan mutant formların seçilmesidir (Dokuzo uz, 1964).
21 tarihinde çıkarılan Ba cılık Yönetmeli i ile sertifikalı fidanla ba tesisi zorunlu kılınmakta ve bu tür çalı maların önemi daha da artmaktadır. Ayrıca Avrupa Birli inin ilgili yönetmeliklerinde belirtildi i gibi üzüm çe itlerinin ticaretinde çe idin ismine do ru olma ko ulu getirilmekte ve gerek duyuldu unda bunun kontrolü talep edilmektedir. Bu amaçla ihtiyaç duyuldu unda güvenilir bir yöntem olan DNA parmak izi analizi kullanılabilecektir. Bu sebeple üzüm çe itlerinin SSR veri bankalarının olu turulması önem kazanmaktadır. Ayrıca yerel asma populasyonuna ait çe it ve klonların tescil a amalarında güvenilir yöntemlerle yapılan genetik tanımlamalar morfolojik tanımlamaları hem teyit edecek hem de morfolojik tanımlamanın bazı olumsuz olabilecek taraflarını elemine edebilecektir. Bu ara tırmada, Manisa Ba cılık Ara tırma Enstitüsü nde klon seleksiyonu çalı maları yoluyla seçilen verim ve kalite yönünden üstün özellikteki klonlar ve tipler ile Ege Bölgesinde yaygın olarak yeti tirilen, ülkemiz için hem ticari hem de gen kayna ı olarak önem ta ıyabilecek olan yerel çe itlerimizde akrabalık ili kilerinin ve DNA kimliklerinin, SSR markörleri yardımıyla ortaya konulması amaçlanmı tır. Çalı mada 2 adet referans çe it olmak üzere toplam 31 tip, klon ve yerel üzüm çe idinin genetik tanımlamaları yapılmı tır.
22 6 2. ÖNCEK ÇALI MALAR 2.1. Bitkilerin Tanımlanmasında Kullanılan Markör Tipleri Kalıtım ekilleri, morfolojik (çiçek rengi), biyokimyasal (izoenzimler) ve DNA düzeyinde (moleküler markörler) izlenebilen karakterlere genetik markörler denir. Genetik markörler gen veya genin bir parçası ya da genler arasındaki bir DNA dizilimidir. Bir tür içerisinde bireyler arasındaki farklılı ı gösteren bir i arettir. Moleküler markörler ise DNA nın aktif bölgelerinden (genler) veya herhangi bir genetik kodlama fonksiyonuna sahip olmayan DNA dizilerinden geli tirilebilirler (Özcan ve ark., 2001). Kalıtımı kolaylıkla belirlenebilen veya gözlemlenebilen DNA dizilimidir. Genetik markörler; Morfolojik markörler, protein markörleri ve DNA markörleri olarak gruplandırılırlar. Morfolojik markörler dominant özelliktedirler, analizlerin kolay olmasına ra men, sayılarının az olu u, çevre ve di er lokuslardan etkilenmeleri nedeniyle fazlaca kullanılmamaktadırlar. Protein markörleri çabuk, güvenilir ve tekrarlanabilir olmasına ra men en büyük dezavantajları sayıca çok az olmalarıdır (Özcan ve ark., 2001) Asmada Günümüze Kadar Yapılan Tanımlama Çalı maları Ampelografik Tanımlamalar Anadolu, Vitis vinifera L. nın gen merkezi, ba cılık kültürünün ba ladı ı ve çe it zenginli inin en üst seviyede oldu u bir bölgedir. Ba cılık yapılan bütün ülkelerde çe itlerin tanımlanması ve
23 7 sınıflandırılmasına yönelik ampelografik çalı malar de i ik ara tırıcılar tarafından sürdürülmü tür. Viala and Vermorel (1910), çe itli ülkelerde yeti tirilen araplık ve sofralık çe idin ampelografik özelliklerini belirlemi tir. Perold (1927), Avrupa nın yakından tanıdı ı sofralık üzüm çe itlerini incelemi ; Manzo and Tamponi (1979), 100 çe idin ampelografik özelliklerini belirlemi tir. Ülkemizde de de i ik ara tırıcılar tarafından farklı yörelerde ve farklı çe itlerde ampelografik tanımlamalar yapılmı tır (Oraman, 1941; Kısakürek, 1950; Fidan ve ark., 1972; Samancı ve Uslu, 1992; Boz ve Çelik, 1998; Kaplan, 1995; Ecevit ve Kelen, 1999; Kader ve Öztürk, 2005) zoenzim (Biyokimyasal) Tanımlamalar Asmalarda 1976 yılında genetik tanımlama amacıyla ilk olarak izoenzimler kullanılmı tır. Melezleme ile edilen ve birbirine yakın olan çe it ve tiplerin tanımlamasında izoenzim teknikleri yaygın olarak kullanılmı tır (Wolfe, 1976; Weeden et. al., 1988; Parfitt and Arulsekar, 1989; Uzun ve lter, 1993; Söylemezo lu ve ark., 1998; A ao lu ve ark., 1999) DNA Markörleri;. DNA Markörleri; Genetik kaynakların karakterizasyonu, Çe itler arasındaki genetik benzerlik ve uzaklıkların belirlenmesi, Islah programında kullanılacak ebeveynlerin belirlenmesi, Yeni geli tirilen çe itlerin koruma altına alınması (ıslahçı hakları),
24 8 Bilinmeyen çe it veya genotiplerin tanımlanması, Genetik haritalama, Islah süresinin kısaltılması (özellikle çok yıllık bitkilerde, testi uzun ve pahalı olan ve çevre ko ullarından etkilenen ve resesif genle kontrol edilen özellikler), Somaklonal varyasyonun belirlenmesi, ebeveyn belirleme, Allel sayısının ve özelliklerinin belirlenmesi, F1 tanımlama, Mutasyonların belirlenmesi konularında kullanılır (Kafkas, ). Farklı genotiplerde DNA dizili inin farklı olması nedeniyle aynı ba latıcı DNA kullanılması ile farklı DNA üretimleri elde edilir. Bu farklı üretimler genetik markör olarak kullanılmaktadır. A a ıda belirtilen çok de i ik tipte DNA markör tipleri geli tirilmi tir. DNA Markörleri; 1. DNA melezleme markörleri - RFLP Markörleri (Restriction Fragment Length Polymorphism) 2. PCR temelli DNA markörleri - RAPD (Random Amplified Polymorphic DNA) Rastgele ço altılmı DNA farklılı ı - AFLP (Amplified Fragment Length Polymorphism) Ço altılan parça uzunlu u farklılı ı - SSR (Simple Sequence Repeat) Basit dizi tekrarları
25 9 - SCAR (Sequence Characterized Amplified Regions) Dizisi tanımlı ço altılan bölgeler eklinde sıralanabilir. deal bir DNA markörün; kurulu ve uygulama maliyetinin dü ük olması, tehlikeli kimyasallarla çalı mayı gerektirmemesi, otomasyona uygun olması, kodominant olması, yüksek polimorfizm göstermesi, birçok bilgi içermesi, sonuçların tekrarlanabilir olması, analizi kolay ve basit olması istenir. Her bir markör sisteminin avantaj ve dezavantajları vardır. Bu nedenle kullanım potansiyelini de erlendirmek çok önemlidir (Kafkas, ). Çizelge 2.1. de bazı markör tiplerinin kar ıla tırılması verilmi tir. Çizelge 2.1. Moleküler markör tekniklerinin kar ıla tırılması Markör Tipi PCR esaslı Polimorfizm Kalıtım RFLP - Dü ük-orta Kodominant RAPD + Orta-Yüksek Dominant SSR + Yüksek Kodominant ISSR + Yüksek Dominant AFLP + Yüksek Dominant IRAP/REMAP + Yüksek Kodominant Morfolojik - Dü ük Dominant/Resessif/Kodominant Protein/isozyme - Dü ük Kodominant STS + Yüksek Kodominant SNP + Yüksek Kodominant SCARS/CAPS + Yüksek Kodominant
26 10 Sonuç olarak DNA markörleri, Genetik kaynakların karakterizasyonu ile ıslah ve genetik programların daha iyi planlanmasını ve daha az maliyetle yapılmasını sa lamaktadır. Geli tirilen markörler, meyve ve çiçek özellikleri ile ili kili ise, özellikle çok yıllık bitkilerde meyve verinceye kadar 2-10 yıl beklemeden erken seleksiyon yapılmasını sa lamaktadır. Analizi pahalı ve uzun olan karakterler ile ili kili markör ise, zamandan tasarruf ve ekonomik kazanç sa lamaktadır. Islahta erken seleksiyon sonucu, deneme alanından ve i gücünden kazanç, daha fazla bitki ile çalı abilme olana ı, ıslahta ba arı oranının artması, daha kısa sürede amaç ve hedeflere ula ılması gibi avantajları bulunmaktadır (Kafkas, 2006) DNA Markörleriyle Yapılan Tanımlamalar Marasalı (1995), Türkiye için önemli olan 11 üzüm çe idinin RFLP tekni ini kullanarak moleküler tanımlamalarını gerçekle tirmi tir. Ye ve arkada ları (1998), yapmı oldukları çalı mada16 üzüm çe idi ve bu çe itlere ait tipler arasındaki ili kiyi ara tırmak için RAPD tekni ini kullanmı lardır. RAPD tekni i denemeye alınan klonlar arasında ayrım sa layamazken, fenotip olarak birbirine benzer çe itler arasındaki ayrım için uygun bir metot oldu u saptanmı tır. A ao lu ve Ergül (1999), Amasya üzüm çe idi ekotipleri olarak bilinen 8 üzüm çe idinin RAPD-PCR tekni i ile genetik tanımlamalarını yapmı tır. Kullanılan 8 adet primerden ikisinin ekotipler bazında kesin bir ayrım sa ladı ı görülmü tür. Yapılan analizlerde bu ekotiplerin ayrı birer çe it olarak tanımlanabilece i tespit edilmi tir. Ergül ve A ao lu (2001),
27 11 asma fidanı üretiminde kullanılan 3 Amerikan asma anacının (5 BB, 110 R, 140 Rug.), 4 farklı üretim istasyonundan alınan örneklerde olası genetik farklılıklar RAPD tekni i ile ara tırılmı tır. Dört farklı damızlık parseline ait 5BB, 110R örneklerinde herhangi bir polimorfizme rastlanmaz iken, 140 Rug. anacında 3 farklı genotipe (1. genotip: Tekirda -Kalecik, 2. genotip: Manisa, 3. genotip: Salihli) ula ılmı tır. SSR veya mikrosatelitler ökaryotik genomlar boyunca da ılmı bulunan ve ardı ık olarak tekrarlanmakta olan 2-6 nükleotid gruplarından olu maktadır. Mikrosatelitleri çevreleyen DNA dizileri genellikle aynı türün bireyleri arasında korunmu olduklarından, farklı genotiplerde çakı an SSR ların PCR primerleri ile ço altılarak seçimine izin vermektedir. Ardı ık SSR tekrarların sayısındaki farklılık PCR sonucu farklı uzunlukta parça ço altımıyla sonuçlanır. Bu tekrarlar çok yakın tür ve çe itler arasında dahi tekrarlanan ünitelerin sayısında de i ikli e yol açan mutasyonlar nedeni ile oldukça polimorfiktir (Gupta ve ark., 1994; Özcan ve ark. dan, 2001). SSR ları çevreleyen korunmu DNA dizileri primer olarak kullanılarak PCR metodu vasıtasıyla bir lokustaki farklı alleller tespit edilebilir (Özcan ve ark., 2001). Sefc (1997), 51 asma çe idinde mikrosatelit markörleri kullanarak ebeveyn tespit edilmeye çalı ılmı tır. Çalı ma sonucunda Cabernet Sauvignon çe idinin orijini Cabernet Franc ve Sauvignon Blanc melezi eklinde oldu unu tespit etmi tir. Asmanın ye il aksamıyla ilgili özelliklerin de i ken ve tanımlamanın zor olması nedeniyle sadece morfolojik özellikler
28 12 kullanılarak asma anaçlarının tanımlanmalarında zorluklarla kar ıla ılmaktadır. Ayrıca bunları dinlenme döneminde ayırt etmek hemen hemen imkansızdır. 58 anacın yaprak ve kambiyum dokularından DNA izole edilmi ve yedi SSR primeri kullanılarak genetik tanımlaması yapılmı tır. Sonuçta bu tekni in anaç ve çe it tanımlanmasında mükemmel bir metot oldu unu belirtmi lerdir (Lin and Walker, 1998). Ergül ve arkada ları (2002), Kalecik Karası üzüm çe idinin 19 klonunda olası genetik farklılık-benzerlikler AFLP tekni i kullanılarak ara tırılmı tır. ki farklı enzim sistemine (Pst1/ Mse1 ve EcoR1/Mse1) ait 6 primer kombinasyonu kullanılmı tır. Pst1/ Mse1 enzim sistemine ait 3 primer kombinasyonunda klonlar arasında bir farklılık bulunamazken EcoR1/Mse1 enzim kombinasyonuna ait primer çiftleri incelendi inde çe ide ait 19 klon ba lıca 2 gruba ayrılmı tır. 5 klonda 3 primerde de polimorfizme ula ılmı tır Asmada Mikrosatelit (SSR) Markörlerin Geli tirilmesi Asmada SSR markörleri ilk olarak, M.R. Thomas, J.E. Bowers ve K.M. Sefc liderli inde çalı an üç ara tırıcı grubu tarafından geli tirilmi tir (Thomas and Scott, 1993; Bowers et al., 1996, 1999a; Sefc et al., 1999). Asmada akrabalık ili kilerini belirlemek, kimlik tanısında kullanılan DNA profillerinin veribankasını geli tirmek ve Vitis vinifera L.nin gen haritalama çalı malarında SSR markörler yo un olarak kullanılır (Thomas et al., 1994; Bowers et al., 1996; Bowers and Meredith, 1997; Lamboy and Alpha, 1998; Bowers et al., 1999b; Riaz and Meredith, 2000).
29 13 Powell ve arkada ları (1996) ve Weising ve arkada ları (1998), bu çe it markörler arasında STMS (Sequence Tagged Microsatellite Sites) nin, kodominant kalıtım, yüksek tekrarlılık, yüksek bilgi içeri i, güvenirlilik ve allel büyüklü ünü kolay skorlama gibi moleküler markörlerin istenilen bir çok özelli ini bir arada topladı ını belirtmi lerdir. Genetik analizlerde; di er metotlara kıyasla SSR markörleri nispeten hızlı ve kolay analiz edilmeyi sa ladı ından pratik ve en önemli araç haline gelmi tir (Morgante and Oliveri, 1993). Arroyo-Garcia ve Martinez-Zapater (2000), araplık ve sofralık üzüm çe itlerinde 9 yeni SSR lokusu (VMC6G8, VMC6D12, VMC6B11, VMC6F11, VMC6G10, VMC6A8, VMC6C7, VMC6C10 ve VMC6E10) dizayn etmi lerdir. PCR ürünlerinin büyüklü ü bp arasında de i mi, allel sayısının her lokusta 8 den 10 a kadar de i ti i gözlenmi tir. Çe itlerin en az % 70 nin her lokusta heterozigot oldu u tespit edilmi tir. araplık ve sofralık üzümlerde allel frekansları genellikle aynıdır. Her çe it 9 lokus tarafından ayırt edilmi tir. Ara tırmacılar çekirdek genomundan daha dü ük mutasyon oranı gösteren kloroplast genomunda da polimorfik mikrosatelit bölgelerinin bu organizmaların biyolojisi hakkındaki bilginin arttırabilece ini belirtmektedirler. Kloroplast genomunda korunmu dizilerin yüksek derecede olmasından dolayı, türlerde yüksek bir oranda çalı abilecek primerlerin dizaynının mümkün olabilmesine olanak sa lamı lardır. Özellikle, kloroplast mikrosatelitlerin (cpssr) sitoplazmik farklılık,
30 14 plastid kalıtımı çalı maları ve gen akı ını görüntülemek için kullanı lı olabildi i belirtilmektedir. Regner ve arkada ları (2006), SSR veri tabanında bulunan dizilimlerini kullanarak Vitis cinsine ait 19 yeni mikrosatellit lokusu detaylı olarak belirlemi lerdir. Asma genotiplerinin ortaya konmasında VRG markörleri kullanılmı tır. Sadece VRG 1, VRG 2, VRG 4, VRG 7, VRG 9, VRG 10, VRG 15 ve VRG 16 markörleri Mendel da ılımına göre heterezigot allel genler olarak da ılım göstermi lerdir. Di er VRG lokusları, VRG 5, VRG 6, VRG 11, VRG 12, VRG 13 ve VRG 17 multi allel profil gösterirken düzgün bir da ılım göstermemi lerdir. Yüksek derecede de i kenlik gösteren VRG mikrosatellitlerin Pinot Noir çe idi içindeki polimorfizmi ortaya çıkarmak için ba arıyla kullanılabilece i ortaya çıkmı tır. Pinot Noir klonları bu markörler kullanılarak ayırt edilmi tir. Goto-Yamamoto ve arkada ları 2006 yılında yaptıkları çal mada, 9 yeni mikrosatelit markör geli tirmi lerdir. Bu markörler ve 8 bilinen mikrosatelit markörle 2 adet Vitis labrusca, Vitis riparia ve Vitis rotundifolia ile birlikte Japon ve Çin çe itlerini (Vitis vinifera L.) de kapsayan 8 adet do u çe idini, 7 adet batı çe idiyle kar ıla tırmı tır. Hesaplanan fenotipik uzaklıklar (payla ılan allel oranı 1) üzümlerin sınıflandırmasıyla aynı olmu tur. Fenotipik uzaklı a dayanan dendogram do u ve batı çe itlerinin yanında Vitis türlerinin farklılı ını açıkça göstermi tir. Sonuçlar, üzüm çe itlerinin gen kayna ı olan do u çe itlerinin önemini destekler nitelikte bulunmu tur.
31 Asmalarda Mikrosatellit Çalı maları Vitis türleri arasındaki taksonomik ili kinin belirlenmesi ve moleküler tanımlamada daha kesin genotip belirlemesi (Sefc et al., 1999), asma çe itleri arasında genetik varyasyon çalı malarında bir çok farklı moleküler markör tipi kullanılması (Parfitt and Arulsekar, 1989; Collins and Symons, 1999; Mauro et al., 1992). Asmada pedigri analizini içeren mikrosatelit markörlerin uygulanması (Bowers and Meredith, 1997). Ebeveyn analizi (Silvestrony et al., 1997; Sefc et al., 1997; Bowers et al., 1999b), çe it tanımlaması ve diversity (çe itlilik, ayrım) çalı maları (Lopes et al., 1999; Sefc et al., 1999, 2000; Aradhya et al., 2003), haritalama (Doligez et al., 2006) ve sinonim tespiti (Lopes et al., 1999; Ulanovsky et al., 2002) konularında mikrosatelit markörler kullanılmı tır. Genetik Kaynakların Karakterizasyonu çalı maları, genetik kayna ın yapısını anlamak, duplike olan genotiplerin belirlenmesi, orijini bilinmeyen genetik materyallerin belirlenmesi, genetik kayna ın tekrar organizasyonu, ıslah programlarında ve genetik çalı malarda kullanılacak ebeveynlerin belirlenmesi gibi nedenlerden dolayı yapılmaktadır. Bu yüzden de ıslahçıların her zaman önemli çalı ma konularından biri olmu tur. Olu turulan gen bankasının tam olarak amacına uygunlu unu sa lamak için, maksimum genetik farklılıklarla birlikte minimum sayıda aksesyonun olması önemlidir. Bu ikisi, bitki ıslahçılarına etkili bir ara tırma ve tarama avantajı sa lar ve gen bankası muhafazası için sınırlı
32 16 olanakların en iyi ekilde kullanımını sunar. Yüzyıllardır klonal olarak ço altılmı ve orijinleri hakkında çok az ey bilinen birçok asma çe idi koleksiyonlarında bu amaca ula mak için çalı ılmı tır. Çünkü tercih edilen asmalar, tarih içinde insanların göç etmesiyle ta ınmı, genetik farklılık olu mu ve birçok çe idin orijini bilinmez hale gelmi tir. Ayrıca, bilinen ve bilinmeyen sinonimler farklı dillerin konu uldu u yerlere ta ınmı tır. Bazen çe it isimleri farklı ekillerde telaffuz edilmi, bazen de götürüldükleri yerlerde tamamen ismi de i tirilmi tir (Dangl et al., 2001). Avusturya gen kayna ı koleksiyonundan toplam 66 üzüm çe idi ve anaçta, 10 mikrosatelit lokus kullanılarak gen farklılı ı de erleri hesaplanmı tır. Ara tırmada yer alan mikrosatelit markörlerin ayrım gücünü ortaya koymak için genetik farklılık de erleri hesaplanmı tır. Olu turulan dendogramda asmalarda genetik farklılık de erleri çe itlerde arasında; anaçlarda 0.29 ila 0.96 arasında iken her ikisi için bu de er arasında tespit etmi tir (Sefc et al., 1998a). Asma genetik kaynakları koruma ve karakterizasyonu için yılındaki EU projesi (Genres CT96 No81 projesi) çerçevesince, asma genetik kaynaklarında çalı an ıslahçılar ve bilim adamlarına Vitis türleri ve asma çe itleri için OIV tanımlama listesinin tamamen revize edilerek bazı kararlar almı lardır. Konu uzmanlarınca asmaların ayrımında normalde dı görünü e, ölçüm, a ırlık ve duyusal (lezzet, koku) incelemeye dayalı ampelografik tanımlamanın yanında mümkünse DNA veya izoenzim analizlerine de yer verilmesi istenmektedir. Ampelometrik yaprak tanımlamalarının önemli bir rol oynadı ı belirtilirken, son geli meler göz önüne alınarak, bol miktarda ve rastgele da ılmı ökaryotik genomlarda ve kodominant Mendel kalıtımına sahip
33 17 SSR markörler asma tanımlama ve ebeveyn analizi için faydalı bir araç oldu u ifade edilmi tir. Bu kapsamda proje partnerleri 49 çe ide uygulanmı olan 6 poliformik SSR markörünün (VVS2, VVMD5, VVMD7, VVMD27, VrZAG62 ve VrZAG79) kullanımı konusunda fikir birli ine varmı lardır. Ayrıca projenin bir di er çıktısı da Avrupa Vitis SSR markör veritabanı kurulması öngörülmektedir ( Önemli ürün türlerinin bulundu u, genetik çe itlili i muhafaza etmek amacı ta ıyan USDA (The United States Department of Agriculture) Ulusal Gen Bankası nda 40 aksesyondan 41 üzüm çe idi 11 polimorfik mikrosatelit lokusunda ara tırılmı tır. Ara tırmada yer alan tüm asmalar Çekirdeksiz ya da benzer isme sahip olan sofralık üzüm çe itleri olmu tur. Morfolojik karakterlerle birlikte yapılan bu çalı mada, bilinen sinonimler do rulanmı, önceki bilinmeyenler ise ortaya çıkarılmı tır. Literatürde iddia edilen bir sinonim DNA verileri ile çürütülmü tür ve bu verilerle ebeveynleri bilinen bazı çe itlerin ebeveynleri do rulanmı tır. USDA koleksiyonunda iki yanlı isimlendirilen çe idin tanımlanmaları da yapılmı tır. Olu turulan UPGMA da (Unweighted Pair-Group Method Using Arithmetic Means) çe itler; ço unlukla Ortado u çe itleri olan 9 çe it; Thomson Seedless ile morfolojik benzerlik gösteren Rusya ve Afganistan dan 22 aksesyon ve ço unlukla Do u Avrupa araplık üzüm çe itlerini içeren 11 aksesyondan olu an grup olmak üzere 3 e ayrılmı tır (Dangl et al., 2001). Crespan et al., (2003), Yerel talyan asma gen kaynaklarını tanımlamı lar ve farklı co rafik bölgelerde yeti tirilen çe itlerin sinonimlerini ortaya çıkarmı lardır. Ba langıçta yapılan ampelografik
34 18 kar ıla tırmalar sonucu sinonim olabilece i öne sürülen çe itlerin (Prosecco lungo ve Tocai nostrano; Aleatico, Vernaccia di Pergola ve Moscatello nero; Bianchetta trevigiana, Vernassiana, Vernanziana ve Senese; Pedevenda ve Verdise; Malvasia bianca lunga ve Fresia; Ranaccio ve Grenache) ampelografik, ampelometrik, GPI ve PGM izoenzim analizleri, mikrosatelit DNA markörler ve aromatik maddelerin kimyasal analizleri gibi de i ik yöntemlerle karakterizasyonu yapılmı tır. Sonuçta, yapılan analizler birbirini tamamlamı ve her aksesyon grupları için ba langıçta öne sürülen sinonimler do rulanmı tır This ve arkada ları (2004), farklı laboratuvarlarda elde edilmi mikrosatelit profillerin kar ıla tırılmasını yapmak amacıyla, 46 üzüm çe idini 6 lokusta (VVMD5, VVMD7, VVMD27, VVS2, VrZAG62 ve VrZAG79) incelemi lerdir. Yaygın bir ekilde kullanılan ve bu çalı madaki 6 markörün, gelecekteki asma çe it analizi için asgari standart markör seti olarak kabul edilmesi önerilmi ve di er çe itlerin, burada sunulan kodlu referans allellerle tanımlanabilece i belirtilmi tir. Akkak ve arkada ları (2007), dünyadaki toplam meyve üretiminin en azından %20 sinin sofralık üzüm oldu una göre en yaygın yeti tirilen meyvelerden birisi asmadır. Bazı üzüm çe itlerimiz uluslararası öneme sahiptir. En önemli çe itler arasında yer alan Cardinal 1939 yılında Flame Tokay (sin. Ahmer Bou Amer) Kaliforniya da Riber (sin. Alphonse Lavallèe) melezlemesiyle elde edilmi tir. Cezayir ve di er Akdeniz ülkelerindeki koleksiyonlardaki asma çe itlerinin DNA ları incelendi inde, a ırtıcı ekilde Cardinal in bu melezlemeden olu madı ı belirtilmi tir. Cardinal in Flame Tokay ın ebeveyni olmadı ı
35 19 bu genetik çalı mada kanıtlanmı tır. Aynı zamanda Ahmer Bou Amer le sinonim oldu u dü ünülen Flame Tokay ın VVS5 mikrosatelit lokusuna göre mutantı oldu u bu çalı mada belirtilmektedir. San Michele all Adige Enstitüsü nde yapılmı olan ıslah çalı malarında elde edilen Incroci Rigotti (IR) melezlerinin ve olası ebeveynleri arasındaki genetik ili kinin belirlenmesi için yapılan çalı mada fluoresan tabanlı kapilar elektroforez teknolojisi ve parçaçık büyüklü ü yazılım programı kullanılmı tır. 11 melez çe itte ve varsayılan 13 ebeveyn çe itler üzerinde 8 SSR markörüyle genotip belirlenmi ve renkli çe itlerin ileri karakterizasyon çalı malarında, IR ve ilgili çe itlerin antosiyanin paterni de elde edilerek de tanımlama yapılmı tır. Sonuçta IR ve IR (Rebo) melezleri aynı bulunmu tur. Aynı çe ide ait aksesyonlar arasında herhangi bir polimorfizm gözlenmemesine ra men. tüm ebeveyn ve IR çe itleri e siz genotipler oldu u ortaya çıkmı tır. Bazı durumlarda, ıslahçılar tarafından belirtilen ebeveynler, IR nin tüm genotipleriyle tutarlılık göstermemi tir (Malossini et al., 2000). Crespan ve Milani (2001) tarafından misket çe itlerinde yapılan çalı mada, morfolojik görünü, tane rengi ve büyüklü ü, olgunla ma zamanı, sofralık ve/veya araplık üretim için e ilimi göz önüne alınarak Misket familyasının çe itlili ini temsil eden. 64 aksesyon analiz edilmi tir. Analizde ayrıca 2 izoenzim ve 25 mikrosatelit lokus incelenmi tir. 64 aksesyondan 20 si moleküler düzeyde birbirinden ayırt edilebilmi, geriye kalan 44 genotipin sinonim oldu u bulunmu tur. Beyaz misket grubunda kırmızı ve pembe taneli üç mutant çe it ayırt
36 20 edilmi tir. Ayrıca, Beyaz misket ve skenderiye Misketi nin Misket ailesinin atası olabilece i belirtilmi tir. spanya da Garnacha, Garnacha Tintoera adıyla yeti tirilen ve bunun yanında sinonimleri olan di er tenturiye aksesyonların 24 mikrosatelit lokusta ve AFLP makörlerle moleküler analizi yapılmı tır (Cabezas et al., 2003). Ara tırma sonuçlarına göre, Garnacha Tinta (kırmızı), Garnacha Gris (gri), Garnacha Blanca (beyaz) ve Garnacha Peluda (tüylü) nın aynı genotipe kar ılık geldi i ve muhtemelen sürekli meydana gelen somaklonal mutasyonlar sonucunda olu tukları dü ünülmektedir. Bunun tersine Garnacha Tintoera, 3 farklı tenturiye genotipi içeren heterojen bir grup oldu u tanımlanmı tır. Fatahi et al., (2003), ran ve ABD den alınan 62 asma (Vitis spp.) aksesyonlarını, kapilar elektroforez parçaçık büyüklü ü sistemi ve fluoresan i aretli primer kullanarak 9 yüksek düzeyde polimorfik mikrosatelit lokusta tanımlamı lardır. Her lokusta gözlenen allel sayıları 4 16 ve heterozigotluk oranı arasında de i mi tir. Genetik benzerlik, her aksesyonun di eriyle kar ıla tırılarak payla ılan allel oranı ile de erlendirilmi tir. Genetik farklılık de erlerinden bir dendogram olu turulmu ve sofralık, araplık ve anaç olmak üzere 3 grup ortaya çıkmı tır. Aynı zamanda bu dendogram, ran sofralık üzüm çe itleri arasında sinonim ve homonimlerin yanısıra 3 klonal grubu (Askari, Bidane ve Yaghoti) da ortaya çıkartmı tır. Kafkasya geçi Bölgesi ve Anadolu dan alınan çe itlerin mikrosatelit tanımlamasının yapıldı ı bu çalı mada 12 mikrosatelit
37 21 markör (VVMD5, VVMD7, VVMD24, VVMD28, VVMD31, VVMD32, VrZAG62, VrZAG79, VVS2, VMC2C3, VMC2H4, VMC5A1) kullanılmı tır. Ermenistan dan 13, Gürcistan dan 41 ve Türkiye den 62 olmak üzere toplam 116 aksesyon üzerinde çalı ılmı tır. Her lokustaki allel sayıları 6 dan (VVMD24) 16 ya (VVMD28 ve VVMD32) kadar de i mi ve 11,9 ortalama göstermi tir. Gözlenen heterozigotluk oranı 0,796 bulunmu tur. Kullanılan markörler içinde en bilgi verici lokusun VMC2H4 ve en az bilgi verenin ise VVMD24 lokusunun oldu u görülmü tür. Türk çe itlerinden Dımı kı, Luvanek, Morek, Sungurlu ve Vilki çe itlerinde 3 allelli durum gözlenmi tir. Çalı mada 20 sinonim ve 6 homonim duruma rastlanmı ve sonuçta 89 faklı üzüm çe idi belirlenmi tir (Vouillamoz et al. 2006). Arroyo-Garcia ve arkada ları (2006), yakla ık 10 ülkeden toplayıp 1000 in üzerinde kültür ve yabani çe itlerinde kloroplast SSR ile yaptıkları çalı malarda üzümün iki orijininden birinin Anadolu di erinin ise spanya oldu u ( araplık çe itler) oldu unu tespit etmi lerdir. Ticari olarak kabul edilen 18 üzüm çe idinin mikrosatelit markörler kullanarak genotip profilleri çıkarılmı ve mevcut referans verilerle kıyasladıklarında; 11 üzüm çe idinin ismine do ru oldu unu 4 üzüm çe idinin ise farklı isimlerle piyasaya arz edildi ini ve kuru üzüm örneklerinin de Sultani çe idi oldu unu, ayrıca Greek Rosaki olarak piyasada bulunan üzüm çe idinin genetik profillerinin incelenmesi sonucu Regina üzüm çe idi oldu unu ortaya çıkartılmı tır. Ayrıca ara tırma sonuçları, DNA düzeyindeki genetik tanımlama metotlarının,
38 22 ya üzümler ve kuru üzümlerde de ba arıyla uygulanabildi i saptanmı tır (Sefc et al., 1998b). Karaa aç (2006), tarafından, Gaziantep iline ait toplam 48 üzüm çe idinin (Vitis vinifera L.) 17 mikrosatelit markör kullanarak genetik düzeyde allel profilleriyle genetik tanımlamaları yapılmı ve aralarındaki genetik benzerlikleri ara tırılmı tır. Lokuslarda gözlenen allel sayısının 13 ile 4 arasında (VVS2, VMC6F1) de i ti ini, beklenen ve gözlenen heterozigotluk oranı ortalama sırasıyla ve olarak bildirilmi tir. Dendogramda 2 ana grup ortaya çıkmı tır. Genotipler arasında Gaziantep ilinden alınan Dusuzu ile Dımı kı çe idi sinonim bulunmu tur. Yine Tekirda ve Gaziantep ilinden alınan Rumi çe itleri dı ında 5 homonim durumuna rastlanmı tır. Gaziantep ten alınan Kı üzümü ve Sergi karası, Sarı kabarcık ve Serpenekıran arasında yakın bir benzerlik oranı gözlenmi tir. Bunların aynı genotipe sahip ve muhtemel somaklonal varyant olabilece i belirtilmektedir. Günümüzde Türkiye asmanın evriminde, üzümün do up geli ti i bölgelere co rafik yakınlı ından dolayı önemli bir rol oynamaktadır. Türkiye de bulunan zengin asma gen kayna ına ra men yeterli genetik analiz yapılmamı tır. elli ve arkada ları (2007) yaptıkları çalı mada, Dimrit ve Gemre üzüm gruplarından olu an 31 üzüm çe idini 8 SSR lokusu ile (VVS2, VVMD5, VVMD7, VVMD24, VVMD27, VVMD28, VrZAG62, VrZAG79) analiz etmi lerdir. SSR verileri üzerinden dendogramda bu iki grup arasındaki genetik farklılık tespit edilmi tir. Bu çalı mada birçok ampelografik özellikleri de incelemi lerdir. Bununla birlikte genotipler ve ekoco rafik da ılım arasındaki genetik ili ki
39 23 açısından önemli bir korelasyon görülmemi tir. Genetik tanımlamada iki Gemre aksesyonu benzer olarak belirlenmi tir. Buna ek olarak bazı sinonim ve homonim genotipler belirlenmi tir. Gizella ve arkada ları (2007), asmanın genomunu ara tırdıkları çalı malarında, 48 üzüm çe idinin 4 enzim sistemi (CO, GOT, AcP, PER) ve 6 mikrosatellite lokusta (VVS2, VVS16, VVMD7, VMC4A1, VMC4G6, VrZaG79) analizini yapmı lardır. Ara tırmada yer alan enzim sonuçlarının tekrarlanabilir oldu u ve odunsu sürgünün örnek alma zamanı asmanın dinlenme periyoduna ba lı kalınmaksızın alınabilece ini belirtmi lerdir. Temel alınan 4 enzimin izoenzim bant desenleri baz alınarak, Badacsony ve Pécs teki FVM Ba cılık ve arapçılık Ara tırma Enstitüsündeki koleksiyon ba ında ara tırılan 48 çe idin ço unun (40 çe it) kimli i belirlenmi tir. zoenzim bant desenleri arasında bir korelasyon bulunmu ve convarietas grubuna ait olan convarietas pontica çe itleri di er convarietasın do u ve batı çe itlerinden farklılık gösterirken, son iki grup birbirinden büyük farklılık göstermemektedir. Pontican çe itleri AcP izoenzimiyle tanımlaması yapılabilmi tir, Fakat nadiren di er iki grupta da görünmü tür. Ara tırmada yer alan 48 çe itten 46 sı mikrosatelit analizlerle tanımlanabilmi tir. Mikrosatelit analizler sonucunda VMC4A1 lokusunda Pinot conculta nın farklılık gösteren 2 çe idinin Pinot gris ve Pinot blanc oldu u belirtilmi tir çi (2007) yaptı ı çalı mada, hastalıklara dayanım ve di er morfolojik özellikler bakımından açılım gösteren F1 popülasyonundan seçilen 60 adet F1 bitkisi ile melezleme için kullanılan talia ve Mercan üzüm çe itleri analiz edilmi tir. Elde edilen amplifikasyon ürünleri 20
40 24 adet SSR primeri için gümü boyama, 24 adet AFLP primeri için radyoaktif i aretleme sonucunda var ve yok seklinde de erlendirildikten sonra 2 testleri sonucunda 1:1 ve 3:1 da ılım gösteren polimorfik lokuslar tespit edilmi tir. Haritalamanın yapılmasında Mapmaker/Exp 3.0 paket programında 3.0 LOD de eri kullanılmı tır. Ana ve babaya ait 2 ayrı genetik ba lantı haritası bulunmu tur. Harita anaya ait 6, babaya ait 1 ba lantı grubu içermektedir. Ba lantı gruplarına yerle en lokusların incelenen hastalıklara dayanım ve morfolojik karakterlere olan ba lantıları regresyon ve varyans analizleri yapılarak belirlenmi tir. Üzüm çe itleri ürün verimi, alkol oranı, asitlik ve antosiyanin de erleri yönünden aralarında farklılık gösterirler. Bu özellikler arap kalitesiyle do rudan ili kili oldu u için, ırada ve araplarda üzüm çe itlerinin do ru bir ekilde tanımlanabilmesi ve belirlenebilmesi son derece önemlidir. Mikrosatelit çalı maları bu tanımlamalarda da önem ta ımaktadır. Faria ve arkada ları (2000), çe it ıralarının ismine do rulu unu DNA tabanlı mikrosatelit yöntemi ile ara tırmı lardır. En önemli 5 porto arabı çe idi (Tinta Roriz, Tinto Cão, Touriga Francesa, Touriga Nacional ve Tinta Barroca) 4 mikrosatelit lokusta çalı ılmı ve ayrıca bu 5 çe it ırası ve bunların kombinasyonu olan 26 ıra karı ımı da 4 mikrosatelit lokusta (VVMD5, VVMD6, VVMD7, VVS2) incelenmi tir. lgili çe itlerin yaprak ve ıralarının bant profilleri arasında bir fark bulunmamı tır. Tüm ıra kombinasyonları çe ide ait bant profilleri bile enlerinin toplamına ba lı olarak beklenen bant
41 25 profillerini göstermi tir. 26 ıra karı ımından 8 i 4 lokusta ayırt edilebilmi tir. Bu metot, sadece belli bir çe idin varlı ı veya yoklu unu belirlemekle kalmamı, aynı zamanda ıranın içinde mevcut olan çe itlerin tespitini de yapmı tır. Balerias-Couto ve Eiras-Dias (2006), yaptıkları çalı mada Touriga Franca, Fernão Pires, Tinta Barroca, Tinta Cão ve Marselan isimli be çe itten üretilen ıra ve araplarda DNA analizlerini baz alınarak moleküler metotlarla tanımlamalar yapılmı tır. Ba langıç olarak ıra ve araptan DNA izolasyonu için üç DNA ekstraksiyon metodu kıyaslanmı ve optimize edilmi tir. Altı nükleer ve iki kloroplast mikrosatelit markörü tek çe itten üretilen araplar ve laboratuarda iki çe idin karı ımıyla hazırlanan ıraların kimlik belirlenmesi için kullanılmı tır. lk de erlendirmeye göre karı ımdaki her çe idin oranı arasındaki olası ili ki ve otomatik dizi analiz sisteminde elde edilen allellerin yo unlu u belirlenmi ; karı ımdaki çe itlerin belirlemenin mümkün olabilece i desteklenmektedir. Ara tırmanın sonuçları araptan DNA izole edilebilece ini ve mikrosatelit amplifikasyonuna uygun oldu unu göstermektedir. Kloroplast genomunda hedefe yönelik kısa DNA parçacıklarının markör olarak ilk kez araplarda asma DNA sını belirlemek için faydalı bir araç oldu unu gösterilmi ; buna göre araplarda orijin sertifikasyonu ve arap kalitesini kontrol etmede bu tekniklerin uygulamasının mümkün oldu u belirtilmektedir SSR Tekni inin Klon ve Tiplerin Tanımlanmasında Kullanımı
42 26 Üzüm ticaretinde Avrupa Birli i Ülkeleri (Reg. 1730/87, Eun 163, 23/6/87) tarafından belirlenen kurallara göre çe idin ismine do ru oldu unun tespiti gerekebilmektedir. Üzüm çe itlerini tanımlamak için mikrosatelit-ssr uygun bir metottur (Sefc et al., 1998). Mikrosatelit (SSR) markörler tekrarlanabilir bantlar olu turması nedeniyle çe itlerin tanımlanmasında tercih edilmektedir (Regner et al.,2000). Klonların korunması, ıslah kurulu larının kanuni haklar elde etmesi ve klonlar arasındaki farklılıkları karakterize etmek ve tanımlamak için biyokimyasal analizler ve ampelografik tanımlamalar yeterli olmamaktadır (Regner et al., 2000). Mikrosatelit-SSR markörler; yüksek stabilite göstermelerinden dolayı klonal asma materyalinin karakterizasyonu ile ilgili verileri saptamak için uygun tekniklerden birisi olarak gösterilmektedir (Regner et al., 2001). Merdinoglu ve arkada ları (2000), tarafından yapılan çalı mada, Vitis vinifera nın 12 çe idine ait 21 klonun testinde üç farklı DNA markörü (RAPD, AFLP, SSR) kullanılmı tır. Her çe idin kendine özgü verdi i bantlar ile çe itlerin ayrımı sa lanmı ve bir dendogram olu turulmu tur. Bu dendogramda 7 grup belirlenmi tir. Çe itler arasındaki genomik benzerli i belirlemek amacıyla yapılan bir çalı mada, Alplerin kuzey ve güney yamaçlarından alınan Schiave grubuna ait 10 üzüm çe idinde AFLP ve SSR kombine olarak kullanılmı tır (Fossati et al., 2001). Sonuç olarak, çe itler arasındaki genetik ili kiyi belirlemek amacıyla yapılan bu çalı mada, AFLP ve SSR ın e it ekilde etkili oldu u belirlenmi tir.
43 27 Silvestroni ve arkada ları (1997), Fortana klonlarında 6 mikrosatelit primerleri ile tekrarlanabilen analizler yapmı lardır. Bütün klonlarda bir primer çifti benzer DNA profilleri gösterirken, 5 tanesi klonlar arasında polimorfizm oldu unu göstermi tir. Ayrıca her klonun allel büyüklüklerini de saptamı lardır. Regner ve arkada ları (2000), Beyaz Riesling üzüm çe idinin 10 klonunda RAPD, SSR ve ISSR markörleriyle genetik poliformizmi ara tırmı lardır. Klonların hepsinde bu üç markör sistemleriyle polimorfizm oldu u görülmü tür. Tanımlamalarda RAPD profilinde stabilite yetersiz görülürken SSR ve ISSR allelleri aynı klon örnekleri içinde tekrarlanabilmi lerdir. SSR ve ISSR markörleri yüksek stabilite göstermelerinden dolayı bu metotlar veri bankası olu turulması için uygundur. Regner ve arkada ları (2001), çalı malarında, Vitis türlerinden alınan 1200 asma genotipinin belirlenmesinde SSR, ISSR, AFLP ve RAPD markörleri kullanılarak yapılmı tır. Polimorfik 6 markör lokusu ile tüm asma çe itlerini ayrılabildiklerini belirten ara tırıcılar, bazı üzüm çe itlerinin orijinini belirlemi lerdir. Klonları ayırt etmede RAPD ve ISSR markörleri uygulanmı tır. Klonları tanımlamanın SSR markörleri ve null allelleriyle de mümkün oldu unu belirtmi lerdir. RFLP ve RAPD markörlerin genetik tanımlamada sınırlı bir kullanımı varken SSR markörleri u anda da güçlü bir araç oldu unu ve RFLP ve RAPD e göre avantajlara sahip oldu unu belirtmi lerdir. Riaz ve arkada ları (2001), genetik açıdan klonların farklılı ı mikrosatelit markörlerle ara tırılmı tır. 22 Pinot Noir ve 22 Chardonnay klonunu ekonomik önemleri ve farklı co rafik orijinlere sahip olmalarından dolayı çalı mada kullanmı lardır. Her bir klon için toplam
44 28 92 mikrosatelit markör kullanılmı, 92 markörden 8 i Pinot Noir ın, 4 ü Chardonnay in klonlarında polimorfizm göstermi lerdir. Çalı mada kullanılan grup içerisinde 7 Pinot Noir ve 5 Chardonnay klonu ayırt edilmi tir. Asma yeti tiricili inde önemli bir yatırımdan önce fidanlıklarda çok genç bitkilerde klonların adına do rulu unu belirlemek için mikrosatelit markörler kullanılabilece ini ve daha çok sayıda polimorfik mikrosatellit markör kullanıldı ında ekonomik öneme sahip Pinot Noir ve Chardonnay klonlarının bireysel ayrımının mümkün oldu unu belirtilmi lerdir. Vignani et al. (2002), 8 mikrosatelit markör kullanarak Sangiovese nin farklı klonlarını ayırt etmi lerdir. Ayrıca, AFLP tekni i de uygulanmı ve sonuçların mikrosatelit lokuslardan elde edilen verilerle örtü tü ü görülmü tür. Islah çalı malarında genetik analizler, klonlar arasındaki farklılı ı ortaya koymada kullanılan bir araçtır. Ço u ba yeti tiricisi geleneksel bir çe idin klonunu üretmeyi tercih eder. Fransa da ve kontrollü üretim yapılan di er ülkelerde ba cılık, AOC etiketi ta ıyan klonlarla üretilme zorunlulu u bulunmaktadır. Dolayısıyla klon materyalinin tanımlanması klonal ıslahta kabul edilen bir yöntemdir. RAPD, AFLP, ISSR ve MSAP (Methylation-Sensitive Amplified Polymorphism) (Imazio et al. 2002) markörleri yüksek derecede polimorfizm verseler bile, SSR markörlerinin stabil, üretken ve genomda bilinen bir pozisyonda olmalarından dolayı daha fazla kabul görmektedirler (Adam-Blondon et al. 2004). Klonal tanımlama amacıyla, VRGs gibi SSR markörleri de oldukça de i ken bölgelere ba lanabildiklerinden, oldukça faydalı bir yöntem olarak ortaya çıkmaktadır (Regner et. al., 2006).
45 29 3. MATERYAL VE YÖNTEM Bu çalı ma, yılları arasında Manisa Ba cılık Ara tırma Enstitüsü Ba ları, Ege Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümü ne ait Biyoteknoloji Laboratuvarı nda yapılmı tır Materyal Ara tırmada kullanılan materyaller, Ba cılık Ara tırma Enstitüsü nde mevcut klon seleksiyonu sonucu elde edilen klonlar, Klon Ba ları, Sultani Çekirdeksiz çe idine ait farklı tipler, Tip Koleksiyon Ba ı ve Ege Bölgesinde yeti tirilen yöresel çe itler de Çe it Koleksiyon Ba larından elde edilmi tir. Ara tırmada, Ege Bölgesi üzüm çe itlerine ait 9 klon (Gemre üzüm çe idinin 6, 11, 12, no lu, Osmanca üzüm çe idinin 26, 39, 40 no lu, pek üzüm çe idinin 4, 13, 25 no lu), 5 tip (Sultani üzüm çe idinin T3, T15, Y3, S4, S6 no lu), 15 yerli çe it (Kozak Beyazı, Kozak Siyahı, Kokulu Misket, Buca Razakısı, Aydın Razakısı, Dumanlı Gemre (kol.) Dumanlı Gemre (os.), Siyah Gemre, Sultani Gemre, Gökçe Gemre, Halis Gemre, Foça Karası (09) Foça Karası (17), Foça Karası (M), Bornova Misketi) ve 2 adet referans (Cabernet Sauvignon ve Merlot) olmak üzere toplam 31 üzüm çe idi kullanılmı tır. Çalı ılan 31 genotipin kısa ampelografik özellikleri Çizelge 3.1. de verilmi tir. Çalı mada adı geçen genotiplerin asmalarından süren genç yaz sürgünlerinin uçları çalı ma materyalini olu turmu tur.
46 30 Çizelge 3.1 Ara tırmada kullanılan üzüm çe itlerine ili kin kısa ampelografik özellikler No Çe it Adı Sinonim Salkım ekli Salkım A ırlı ı (g) Tane ekli Tane Rengi Tane A ırlı ı (g) Aroma De. ekli Verim (kg/da) Olgunla ma 1 Kozak Beyazı 2 Kozak Siyahı 3 Kokulu Misket 4 Bornova Misketi 5 Buca Razakısı 6 Aydın Razakısı ika Konik Orta ( ) Yumurta Ye il-sarı ri ( ) Tatlı, hafif mayho - Da ınık konik, dallı Sofralık Orta ( ) Basık oval Siyah Orta (4-5) Tatlı Sofralık - Dallı konik Orta (406) Yuvarlak Ye il-sarı Orta (2.8) Hafif misket - Konik-silindirik Orta ( ) Yuvarlak Ye il-sarı Küçük (2.0) Konik ri (550) Uzun elips Ye il-sarı Dallı konik Orta (488) Kısa elips Ye il-sarı Ç. büyük (8.0) Ç. büyük (7.0) Misket kokulu ıralık araplık Az tatlı Sofralık Fazla (1797) Fazla (2806) Ç. fazla (3813) Ç. Fazla (3591) Fazla (1692) Nötral Sofralık Orta (1498) 7 Foça Karası (09) 2 Konik Orta (360) G. yumurtamsı Kırmızı-siyah Orta (3.7) Nötral araplık Orta (1470) Geç Geç Orta-geç Orta erken Geç Geç Orta mevsim 8 Foça Karası (17) 2 Kanatlı konik Orta (370) Kısa elips Kırmızı-siyah Orta (3.0) Nötral araplık Az (958) Orta mevsim 9 Foça Karası (M) 2 Silindirik Küçük (227) G. yumurtamsı Pembe Orta (3.4) Nötral araplık Orta (1453) Orta mevsim 10 Osmanca (26) 5 Osmancık Konik Orta (500) Yuvarlak Ye il-sarı Orta (3.7) Nötral Sofralık Ç. Fazla (2790) Geç 1 Anonim, Dilli, Çelik, Samancı ve Uslu, Kader, 2005
47 31 Çizelge 3.1 (devamı) No Çe it Adı Sinonim 11 Osmanca (39) 12 Osmanca (40) 5 5 Salkım ekli Salkım A ırlı ı (g) Tane ekli Tane Rengi Tane a ırlı ı (g) Aroma Osmancık Konik Orta (520) Yuvarlak Ye il-sarı Orta (4.0) Nötral Sofralık Osmancık Konik Orta (500) Yuvarlak Ye il-sarı Orta (3.6) Nötral Sofralık 13 pek (4) 2 Pek Dallı konik Orta (545) Kısa Elips Ye il Orta (3,9) Nötral Sofralık De. ekli Verim (kg/da) Ç.fazla (3200) Ç. fazla (2740) Fazla (2078) Olgunla ma Geç Geç Geç 14 pek (13) 2 Pek Dallı konik Orta (499) Kısa Elips Ye il Fazla (4,6) Az tatlı Sofralık Az (946) Geç 15 pek (25) 2 Pek Dallı konik Orta (371) Kısa Elips Ye il-sarı Fazla (4,3) Az tatlı Sofralık 16 Sultani Çek. T3 17 Sultani Çek. T15 18 Sultani Çek. Y3 19 Sultani Çek. S4 20 Sultani Çek. S6 6 Kanatlı uzun silindirik 6 Kanatlı uzun silindirik 6 Kanatlı uzun silindirik 6 Kanatlı uzun silindirik 6 Kanatlı uzun silindirik Orta (370) Yumurta, basık yumurta Ye il-sarı Ç. az (1.3) Nötral Kurutmalık Orta (429) Basık yumurta Ye il-sarı Ç. az (1.4) Nötral Kurutmalık Orta (326) Yumurta, basık yumurta Ye il-sarı Ç. az (1.4) Nötral Kurutmalık Orta (329) Basık yumurta Ye il-sarı Az (1.6) Nötral Sofralık Orta (494) Yumurta Ye il-sarı Az (1.8) Nötral Sofralık Orta (1376) Fazla (2280) Ç. fazla (2780) Fazla (1950) Fazla (1800) Fazla 82540) Geç Orta mevsim Orta mevsim Orta mevsim Orta mevsim Orta mevsim 6 Ilgın vd.,1999
48 32 Çizelge 3.1 (devamı) No Çe it Adı Sinonim Salkım ekli Salkım A ırlı ı (g) Tane ekli Tane Rengi Tane a ırlı ı (g) Aroma De. ekli 21 Pembe Gemre (6) 7 Dallı konik ri (739) Yuvarlak Pembe Ç. fazla (7.5) Nötral Sofralık 22 Pembe Gemre (11) 7 Dallı konik ri (755) Yuvarlak Pembe Ç. fazla (7.0) Nötral Sofralık 23 Pembe Gemre (12) 7 Dallı konik ri (801) Yuvarlak Pembe Ç. fazla (7.2) Nötral Sofralık 24 Dumanlı Gemre (kol.) 7 Konik ri (551) Yuvarlak K. kırmızı-mor Orta (3.5) Az tatlı Sofralık 25 Dumanlı Gemre (os.) 2 Dallı da ınık konikı Orta (362) Yuvarlak K. kırmızı-mor Orta (4.0) Hafif misket Sofralık 26 Siyah Gemre 27 Sultani Gemre 7 7 Dallı da ınık konik Orta (431) Yuvarlak Kırmızı-siyah Orta (2.8) Az tatlı Konik Orta (426) Kısa elips, yumurta Sofralık- ıralık K. kırmızı-mor Orta (3.5) Nötral Sofralık 28 Gökçe Gemre 7 Dallı konik Orta (507) Yuvarlak K. kırmızı-mor Orta (3.1) Nötral Sofralık 29 Halis Gemre 7 Konik Orta (471) Yuvarlak K. kırmızı-mor Orta (3.1) Az tatlı Sofralık 7 Dilli ve Kader, 2005 Verim (kg/da) Fazla (2397) Fazla (2865) Fazla (2569) Ç.fazla (3937) Fazla (1968) Fazla (2805) Ç. fazla (3038) Fazla (2606) Fazla (2320) Olgunla ma Geç Geç Geç Geç Geç Geç Geç Geç Geç




49 33 Kozak Beyazı Kozak Siyahı Kokulu Misket Bornova Misketi ekil Ara tırmada Kullanılan Üzüm Çe itleri


")

50 34 Buca Razakısı Aydın Razakısı Foça Karası (09) Foça Karası (17) ekil (Devamı)



")
51 35 Foça Karası (M) Osmanca (26) ekil (Devamı) Osmanca (39) Osmanca (40)
 pek")
")


52 36 pek (4) pek (13) pek (25) Sultani Çekirdeksiz (T3) ekil (Devamı)

 Sultani")
 ekil 3. 1.")

53 37 Sultani Çekirdeksiz (T15) Sultani Çekirdeksiz (Y3) Sultani Çekirdeksiz (S4) Sultani Çekirdeksiz (S6) ekil (Devamı)
")

")
54 38 Pembe Gemre (6) Pembe Gemre (11) Pembe Gemre (12)


")

55 39 ekil (Devamı) Dumanlı Gemre (Kol) Dumanlı Gemre (Os.)

")
56 40 Siyah Gemre Sultani Gemre ekil (Devamı) Gökçe Gemre Halis Gemre ekil (Devamı)
57 Yöntem Manisa Ba cılık Ara tırma Enstitüsü nde klon seleksiyonu sonucu elde edilmi Çekirdeksiz tiplerin, klonların ve Ege Bölgesi için önemi olan bazı yerel çe itlerimizin DNA düzeyinde benzerlik ve farklılıkların ara tırılması ve tanımlaması için SSR tekni i kullanılmı tır. Bunun için DNA izolasyonu, PCR uygulaması, SSR tekni inin poliakrilamid jel kullanılarak dikey elektroforezde uygulanması ve jel sonuçlarının gümü boyama ile görüntülenmesi ve sonuçların de erlendirilmesi eklinde gerçekle tirilmi tir DNA izolasyonu DNA izolasyonu Lefort et al. (1998) yöntemine göre yapılmı tır. Çe itlerin genç ve yarı genç yapraklarından alınan örneklerle DNA izolasyonu gerçekle mi tir. zolasyonda izlenen a amalar a a ıda belirtilmi tir. Buna göre; Genç yaprak havana konulup sıvı azotla havanda iyice ezildi, Toz haline gelen yaprak örne inden 0,1 gr alınarak 1,5 µl ependorf tüplere aktarıldı, Üzerine 1 ml DNA ekstraksiyon tampon çözeltisi ilave edildi ve homojen hale gelinceye kadar ependorf tüpe hafifçe vurarak karı ması sa landı, 65 o C de 15 dakika sıcak suda bekletildi,
58 42 Üzerine 0,5 ml kloroform/isoamil alkol (24:1) karı ımı eklendi ve hafif bir ekilde iyice çalkalandı, bu arada buzda bekletildi, Oda sıcaklı ında, 1 dakika rpm hızında santrifüj edildi, Ependorf tüpün üst kısmındaki sıvı, di er bir temiz ependorf tüpe aktarıldı, Üzerine 0,8 ml isopropanol eklendi, 15 dakika buz üzerinde tutuldu ve 1 dakika rpm hızında santrifüj edildi, Üst sıvı uzakla tırıldı, 1 ml % 70 lik ethanol eklendi ve yine 1 dakika rpm hızında santrifüj edildi, Eklenen ethanol uzakla tırıldı ve ependorf tüpün a zı açık bırakılarak alkolün tamamen uçması sa landı, DNA, 100 µl TE tampon çözeltisi (ph 8,0) içinde çözüldü, Her 100 µl için 1 µl RNase-A eklenerek, 37 o C de 15 dakika etüvde bekletildi. DNA izolasyonunda kullanılan çözeltiler a a ıda verilmi tir. Ekstraksiyon tampon çözeltisi (100 ml için): 4 ml TRIS (50 mm, ph 8,0) 8 ml EDTA (50 mm, ph 8,0) 20 ml LiCl (4M) 2 g CTAB (% 1) 4 g PVP (% 2)
59 43 1 ml TWEEN 20 ( % 0,5) Kloroform/isoamil alkol; (24:1) (hacim:hacim) TE çözeltisi; 10 mm TRIS-HCl ve 1 mm EDTA (ph 8,0) Rnase-A (Sigma R6513); 10 mg/ml PCR uygulaması PCR metodu basitçe tüp içerisinde nükleik asitlerin uygun ko ullarda ço altılması esasına dayanır. DNA ço altımı için PTC-200 MJ Research Type Thermocycler cihazı ve PCR optimizasyon çalı maları yapılmı tır. PCR ço altma ko ullar 3.5 µl DNA, 2. 5 µl 10X Buffer, 2.5 µl MgCl 2, 0.8 µl dntp, 0.8 µl primer 1 (forward), 0.8 µl primer 2 (reverse), 0.3 µl Taq, 14.8 µl su olmak üzere toplam 25 µl PCR karı ımı hazırlanmı tır (Çizelge 3.2). DNA ço altımı için kullanılan PCR programı C de 3 dakika, C de 1 dakika, C de 1 dakika (sıcaklık primerin ba lanma derecesine ba lı olarak de i ecektir) C de 2 dakika (2., 3. ve 4. a amaları 10 döngü) C de 1 dakika C de 2 dakika C de 2 dakika (5,6,7 a amaları 20 döngü) 8. Son yazılım safhasında 72 C de 10 dakika
60 44 Çizelge 3.2 PCR ço altma ö eleri konsantrasyon ve miktarları PCR Ço altma ö eleri Kullanılan konsantrasyon PCR da kullanılan miktar (µl) DNA 100 ng 3,5 10XBuffer 1X 2,5 MgCl mm 2,5 dntp 2.5 mm 0,8 Primer ng 0,8 Primer ng 0,8 Taq 1.5 U 0,3 Su 13,8 Toplam 25 Çizelge 3.3 Primerlerin PCR daki döngü ko ulları Ön denatüras yon 94 ºC 3 dk Denatüra syon 94 ºC 1 dk Primer ba lanma * 55 ºC 1 dk Yeni iplikçik yazılımı 72 ºC 2 dk Son yazılım 72 ºC 10 dk *: Sıcaklık primerin ba lanma derecesine ba lı olarak de i mi tir. Toplam Döngü sayısı Çalı mada Kullanılan SSR Primerleri Çalı mada, GENRES 081 Avrupa Birli i Ara tırma Projesince, Avrupa daki asma çe it koleksiyonları için kullanılan ve artık tüm dünya tarafından minimum standart set olarak kabul gören VVS2, VVMD5, VVMD7, VVMD27, VrZAG62 ve VrZAG79 mikrosatelit lokusları ve daha önceki çalı malarda informatif oldu u tespit edilen primerler arasından seçilen VVS1, VVS3, VVS4, VVMD6, VVMD17, VVMD24, VVMD28, VVMD31, VrZAG29, VrZAG67 lokusları da dahil olmak üzere olmak üzere toplam 16 SSR primeri
61 45 kullanılmı tır. Her SSR primeri, forward ve reverse eklinde olup primerlere ait baz dizileri Çizelge 3.4 de verilmi tir. Çizelge 3.4 Ara tırmada kullanılan primerlere ait bilgiler Locus Primers 5'-3' Referans VVS1 F R ACAATTGGAAACCGCGTGGAG CTTCTCAATGATATCTAAAACCATG VVS2 VVS3 F R F R CAGCCCGTAAATGTATCCATC AAATTCAAAATTCTAATTCAACTGG TGCCCTATCAATTAGTTCACCTA TCGACTTTGATATATTGATGATT Thomas and Scott 1993 VVS4 F R CCATCAGTGATAAAACCTAATGCC CCCACCTTGCCCTTAGATGTTA VVMD5 F R CTAGAGCTACGCCAATCCAA TATACCAAAAATCATATTCCTAAA Bowers et al ve 1999a VVMD6 F R ATCTCTAACCCTAAAACCAT CTGTGCTAAGACGAAGAAGA Bowers et al VVMD7 F R AGAGTTGCGGAGAACAGGAT CGAACCTTCACACHCTTGAT Bowers et al ve 1999a VVMD17 F R TGA CTC GCC AAA ATC TGA CG CAC ACA TAT CAT CAC CAC ACG G Bowers and Meredith 1997 VVMD24 F R GTG GAT GAT GGA GTA GTC ACG C GAT TTT AGG TTC ATG TTG GTG AAG G VVMD27 VVMD28 F R F R GTA CCA GAT CTG AAT ACA TCC GTA AGT ACG GGT ATA GAG CAA ACG GTG T AACAATTCAATGAAAAGAGAGAGAGAGA TCATCAATTTCGTATCTCTATTTGCTG Bowers et al ve 1999a VVMD31 F R CAGTGGTTTTTCTTAAAGTTTCAAGG CTCTGTGAAAGAGGAAGAGACGC VrZAG29 F R ATA ACC AGG ACA AGT TAT TCA AGC C ACC CAA TTG ACC ATC TTT TAT GCT G VrZAG62 VrZAG67 F R F R GGT GAA ATG GGC ACC GAA CAC ACG C CCA TGT CTC TCC TCA GCT TCT CAG C ACC TGG CCC GAC TCC TCT TGT ATG C TCC TGC CGG CGA TAA CCA AGC TAT G Sefc et al VrZAG79 F R AGATTGTGGAGGAGGGAACAAACCG TGCCCCCATTTTCAAACTCCCTTC C Ara tırma için seçilen 16 SSR (Simple Sequence Repeats) primerleri, MWG-BIOTECH Inc. firması tarafından temin edilmi tir.
62 Agaroz Jel Elektroforezi SSR-PCR tekni i ile elde edilen amplifikasyon ürünleri, poliakrilamid jele yüklemeden önce 10 µl alınarak % 3 lük agaroz jelde kontrol edilmi tir. Agaroz jele yüklenen ürünler, yatay elektoforezde 1X TBE (Tris, Borik asit, EDTA) tampon çözeltisi içinde 100 V da yakla ık 2,5-3 saat yürütüldükten sonra, etidium bromid (0.01 mg/ml) ile boyanarak; UV altında ( =302 nm) gözlemlenmi tir. 9 klon, 5 tip, 15 yerli çe it ve 2 adet referans çe it ve 1 negatif kontrol olmak üzere toplam 32 adet bireyde, 16 adet SSR primerlerinin kullanılmasıyla elde edilip agaroz jelde kontrol edilmi ço altma ürünleri (bantlar), % 6 lık poliakrilamid jelde elektroforez edilmi ir Poliakrilamid Jel Elektroforezi ve Jel Boyama Welsh ve arkada ları (1995), uzunlu u az olan PCR ürünlerinin ayrımında genellikle poliakrilamid jel elektroforezinin (PAGE-jel) kullanıldı ını belirtmi lerdir. SSR tekni i ile genotiplerin moleküler analizi için standart PAGE-jel (%6 lık poliakrilamid, 19:1 akrilamid: bis; 8 M üre, 0,5x TBE (Tris, Borik asit, EDTA) buffer) kullanılmı tır. Poliakrilamid jele yüklemeden önce SSR markörlerine ait PCR ürünleri ve PCR ürünlerinin yarısı kadar yükleme boyası (loading buffer; %98 formamide, 10 mmol/l EDTA, %0,05 bromofenol mavisi ve xylene cyanol) ile 94 C de 5 dakika denatüre edilip, hemen buz üzerine
63 47 alınmı tır. PAGE-jel (poliakrilamid) 55 C ye ula tıktan sonra, denatüre edilmi her bir reaksiyondan 10µl olacak ekilde %6 lık poliakrilamid jele yükleme gerçekle tirilmi ve 100 W da 2,5-3 saat süreyle yürütülmü tür. 38x45 cm boyutundaki dizi analizi jel tankında (dikey elektroforez) yürütülen bantların görüntülenmesi amacıyla ise gümü boyama (silver staining) (Silver Sequence Kit, Promega) uygulanmı tır. Daha sonra tarayıcı vasıtasıyla görüntülenerek bilgisayar ortamına aktarılmı tır. Bio-PROF L Bio-1D++ programında her bir lokusa ait allel büyüklü ü, baz çifti (bp, basepair) olarak analiz edilmi tir Allel De erlerinin Belirlenmesi ve Genetik Analizler Ara tırmada; genetik parametreler olarak her lokusa ait allel sayısı (n), allel frekansı, beklenen (H e ) ve gözlenen heterozigotluk oranı (H o ), tahmin edilen sessiz allel (null) frekansı (r) ve tespit olasılı ı (PI, Probability of Identity).IDENTITY 1.0 (Wagner and Sefc 1999) 2 programı kullanılarak tespit edilmi tir. Beklenen heterozigotluk 1- p i eklinde hesaplanarak genetik farklılık ölçümü yapılmı tır. Buradaki p i de eri, çalı ılan örnekteki i ninci allelin frekansını göstermektedir (Nei 1987). Gözlenen heterozigotluk ise, heterozigot genotiplerle analiz edilen toplam genotip arasındaki orandır. Null allel varlı ının tahmini, (H e - H o )/(1+ H e ) eklinde hesaplanmı tır (Brookfield 1996). Tespit olasılı ı (PI) (Paetkau et al. 1995) iki rastgele seçilmi bireylerin aynı SSR profile sahip olma olasılı ını ifade etmektedir. Bu de er, p 4 i + (2p i p j ) 2 eklinde hesaplanır. Burada p i ve p j, sırasıyla i ve j allellerinin frekansını göstermektedir. Daha sonra benzerlik oranı indeksleri Microsat (Minch et al. 1995) programı ile genetik uzaklık D=1- (payla ılan allel oranı) (benzersizlik oranı, disimilarity) hesaplanarak
64 48 bulunmu tur. Bu de er daha sonra similarity yani benzerlik oranına dönü türülmü tür. Genotiplere ait genetik ili ki dendogramı. UPGMA (Unweighted Pair-Group Method using Arithmetic means) yöntemi kullanılarak NTsys (versiyon 2.02g, Exeter Software, Setauket, NY) programı ile olu turulmu tur.
65 49 4. ARA TIRMA BULGULARI 4.1. DNA zolasyonu DNA izolasyonu sonucunda elde edilen DNA ların, saflık ve miktar de erleri NanoDrop ND1000 ile belirlenmi tir (Çizelge 4.1). DNA ların miktar tayininde absorbsiyon temeline dayanan spektrometrik yöntemler kullanılır. Ölçümlerde nükleik asitler 260 nm de absorbsiyon de eri (A260); oldukça saf olarak izole edilen nükleik asitlerin mikrogram düzeyinde miktarların belirlenmesinde kullanılır. Sadece A260 de eri DNA ve RNA yı tam olarak ayırt etmedi i için proteinlerin maksimum absorbsiyon özelli i gösterdi i 280 nm deki ölçüm de erleri de önemlidir. A260/280 oranı DNA nın saflı ı hakkında bilgi vermektedir. Bu oranının arasında olması istenmektedir. Çizelge 4.1 Ara tırmada kullanılan üzüm çe itlerine ait DNA miktarı ve saflık dereceleri. Genotip no* DNA miktarı (ng/µl) Saflık dereceleri (A260/A280) Genotip no* DNA miktarı (ng/µl) Saflık dereceleri (A260/A280) , , , , , , , , , , , , , , , , , , , , , , , , , , , , ,90 * Çe itlerin numaralandırma sırası Çizelge 3.1 e göre yapılmı tır.
66 50 Bu veriler incelendi inde DNA miktarlarının 1509 ile 5026 ng/ µl arasında de i ti i görülmü tür. En az DNA yo unlu u 16 numaralı Sultani Çek. T3 çe idinde görülürken, 5026 ng/µl ile 22 numaralı Pembe Gemre (11) en fazla DNA yo unlu una sahip çe it olmu tur. DNA saflı ının genellikle arasında olması istenir. Çalı mada elde edilen DNA saflıklarının aralı ında de i ti i Çizelge 4.1 de görülmektedir. Buna göre tüm genotiplerin DNA saflıkları yeterli düzeydedir Allel Büyüklükleri Çalı ılan primerlerde parçacık ço altımı olup olmadı ını anlamak için PCR ürünleri agaroz jele yüklenmi, ço altım gerçekle mi ise poliakrilamid jele örnekler yüklenmi tir. Ara tırmada kullanılan primerler için laboratuarda amplifikasyon standardizasyonu yapılarak örnekler üzerinde ara tırmalar yürütülmü tür. VVMD24 primeri PCR reaksiyonlarındaki optimize çalı malarına ra men sonuç elde edilememi tir. PCR ürünleri ve PCR hacminin yarısı kadar loading buffer olmak üzere toplam 10 µl hacim 94 C de 5 dakika denatüre edilmi ve örnekler poliakrilamid jele yüklenmi tir. Bantların görüntülenebilmesi amacıyla ise gümü boyama uygulanmı tır. Daha sonra tarayıcı vasıtasıyla görüntülenerek bilgisayar ortamına aktarılmı tır. Elde edilen görüntüler Bio-PROF L Bio-1D++ programında analiz edilmi ve her bir lokusa ait allel büyüklükleri baz çifti (bp, basepair) olarak Çizelge 4.2. de gösterilmi tir. Ayrıca iki Fransız çe it Cabernet Sauvignon ve Merlot çe itleri de referans çe it olarak örneklerle beraber analiz edilmi tir.
67 51 Ara tırmada kullanılan 15 primerden, VVMD17 ve VrZAG29 primerlerinin ön istatistik analiz de erleri nedeniyle genetik analizlere dahil edilmemi tir. Yalnızca allell de erleri verilmi tir. Çalı ılan genotiplerde, istatistik analizleri yapılan 13 lokusta toplam 143 allel tespit edilmi tir. Allel büyüklüklerine bakıldı ında, Sultani Çekirdeksiz çe idinin T15 ve Y3 tipleri, pek çe idinin 13 ve 25, Osmanca üzüm çe idinin 26 ve 40 no lu klonları her lokusta ayni allelleri vermi tir. Sultani Çekirdeksiz çe idinin T3, T15 ve Y3 tipleri VVMD6 ve VVMD31 lokuslarında 3 allel; Sultani Çekirdeksiz çe idinin S4 ve S6 tipleri VVMD31 lokusunda 2 allel bakımından farklılık göstermi tir. Osmanca 26, 39, 40 no lu klonları arasında 3 allel farklı bulunmu tur. Pembe Gemre 6 ve 12 no lu klonları sadece VrZAG62 lokusunda 1 allel; Pembe Gemre 6, 11 ve 12 no lu klonları 2 allel tarafından ayrılmı tır. pek 4, 13 ve 25 no lu klonları 2 allel; Buca Razakısı ve Aydın Razakısı arasında ise 5 allel farklı bulunmu tur.
68 52 Çizelge 4.2 Çe it, tip ve klonların 15 lokustaki allel büyüklükleri (bp) 1 Mikrosatelit Lokuslar No ÇE T ADI VVS1 VVS1 VVS2 2 VVS2 2 VVS3 VVS3 VVS4 VVS4 1 Kozak Beyazı Kozak Siyahı Kokulu Misket Bornova Misketi Buca Razakısı Aydın Razakısı Foça Karası (09) Foça Karası (17) Foça Karası (M) Osmanca (26) Osmanca (39) Osmanca (40) pek (4) pek (13) pek (25) Sultani Çek. T Sultani Çek. T Sultani Çek. Y Sultani Çek. S Sultani Çek. S Pembe Gemre Pembe Gemre Pembe Gemre Dumanlı Ge. (kol.) Dumanlı Ge. (os.) Siyah Gemre Sultani Gemre Gökçe Gemre Halis Gemre C. Sauvignon Merlot Referans çe itler koyu ile belirtilmi tir 2 Asma çe it analizi için minimum standart mikrosatelit markör seti olarak kabul edilen markörler (This et al.2004)
69 Çizelge 4.2 nin devamı No ÇE T ADI Mikrosatelit Lokuslar VVMD5 2 VVMD5 2 VVMD6 VVMD6 VVMD7 2 VVMD7 2 1 Kozak Beyazı Kozak Siyahı Kokulu Misket Bornova Misketi Buca Razakısı Aydın Razakısı Foça Karası (09) Foça Karası (17) Foça Karası (M) Osmanca (26) Osmanca (39) Osmanca (40) pek (4) pek (13) pek (25) Sultani Çek. T Sultani Çek. T Sultani Çek. Y Sultani Çek. S Sultani Çek. S Pembe Gemre Pembe Gemre Pembe Gemre Dumanlı Ge. (kol.) Dumanlı Ge. (os.) Siyah Gemre Sultani Gemre Gökçe Gemre Halis Gemre C. Sauvignon Merlot Referans çe itler koyu ile belirtilmi tir 2 Asma çe it analizi için minimum standart mikrosatelit markör seti olarak kabul edilen markörler (This et al.2004) 53
70 54 Çizelge 4.2 nin devamı No ÇE T ADI Mikrosatelit Lokuslar VVMD17 VVMD17 VVMD27 2 VVMD27 2 VVMD28 VVMD28 1 Kozak Beyazı Kozak Siyahı Kokulu Misket Bornova Misketi Buca Razakısı Aydın Razakısı Foça Karası (09) Foça Karası (17) Foça Karası (M) Osmanca (26) Osmanca (39) Osmanca (40) pek (4) pek (13) pek (25) Sultani Çek. T Sultani Çek. T Sultani Çek. Y Sultani Çek. S Sultani Çek. S Pembe Gemre Pembe Gemre Pembe Gemre Dumanlı Ge Dumanlı (k l ) Ge Siyah ( ) Gemre Sultani Gemre Gökçe Gemre Halis Gemre C. Sauvignon Merlot Referans çe itler koyu ile belirtilmi tir 2 Asma çe it analizi için minimum standart mikrosatelit markör seti olarak kabul edilen markörler (This et al.2004)
71 55 Çizelge 4.2 nin devamı N o ÇE T ADI Mikrosatelit Lokuslar VVMD31 VVMD31 VrZAG29 VrZAG29 VrZAG62 2 VrZAG Kozak Beyazı Kozak Siyahı Kokulu Misket Bornova Misketi Buca Razakısı Aydın Razakısı Foça Karası (09) Foça Karası (17) Foça Karası (M) Osmanca (26) Osmanca (39) Osmanca (40) pek (4) pek (13) pek (25) Sultani Çek. T Sultani Çek. T Sultani Çek. Y Sultani Çek. S Sultani Çek. S Pembe Gemre Pembe Gemre Pembe Gemre Dumanlı Ge. (kol.) Dumanlı Ge. (os.) Siyah Gemre Sultani Gemre Gökçe Gemre Halis Gemre C.Sauvignon Merlot : Referans çe itler koyu belirtilmi tir 2:Asma çe it analizi için minimum standart mikrosatelit markör seti olarak kabul edilen markörler (This et al. 2004)
72 56 Çizelge 4.2 nin devamı No ÇE T ADI Mikrosatelit Lokuslar VrZAG67 VrZAG67 VrZAG 2 79 VrZAG Kozak Beyazı Kozak Siyahı Kokulu Misket Bornova Misketi Buca Razakısı Aydın Razakısı Foça Karası (09) Foça Karası (17) Foça Karası (M) Osmanca (26) Osmanca (39) Osmanca (40) pek (4) pek (13) pek (25) Sultani Çek. T Sultani Çek. T Sultani Çek. Y Sultani Çek. S Sultani Çek. S Pembe Gemre Pembe Gemre Pembe Gemre Dumanlı Ge. (kol.) Dumanlı Ge. (os.) Siyah Gemre Sultani Gemre Gökçe Gemre Halis Gemre C.Sauvignon Merlot : Referans çe itler koyu belirtilmi tir 2:Asma çe it analizi için minimum standart mikrosatelit markör seti olarak kabul edilen markörler (This et al. 2004)
73 Yazılım Programları ile Sonuçların De erlendirilmesi 31 asma genotipinin sonuç veren 13 lokusla analizi, çalı ılan lokuslardaki allel sayıları, heterozigotluk oranları, tespit olasılı ı de eri ve sessiz (null) allel frekansı Çizelge 4.3 de sunulmu tur Çizelge 4.3. Çalı ılan lokuslardaki allel sayıları (N), beklenen heterozigotluk (He), gözlenen heterozigotluk (Ho), tespit olasılı ı (PI) de eri ve sessiz (null) allel frekansı Analiz edilen lokuslar Lokusta gözlenen allel sayısı (N) Beklenen heterozigotluk oranı (He) Gözlenen heterozigotluk oranı (Ho) Tespit olasılı ı (PI) de eri Sessiz (null) allel frekansı (r) VVS VVS VVS VVS VVMD VVMD VVMD VVMD VVMD VVMD VrZAG VrZAG VrZAG Toplam Ortalama VVS1, VVS2, VVS3, VVS4, VVMD5, VVMD6, VVMD7, VVMD27, VVMD28, VVMD31, VrZAG62, VrZAG67, VrZAG79 SSR lokusları itibari ile allel sayıları sırası ile 9, 10, 6, 9, 10, 10, 12, 11, 14, 13, 15, 10, 14 olarak tespit edilmi tir. He ve Ho de erleri lokuslara göre de i kenlik göstermi tir. VVS1, VVMD5, VVMD6, VVMD7, VVMD27, VVMD28, VVMD31, VrZAG62, VrZAG67, VrZAG79. lokuslarında He, VVS2,
74 58 VVS3, VVS4.lokuslarında ise Ho de erleri yüksek bulunmu tur. PI de erleri ise 0, arasında de i mi tir. Null allel frekansı ise lokuslarda kabul edilebilir sınırlar içerisinde (genel olarak 0.1 in altında) bulunmu tur. Lokuslar açısından önemli olan di er bir genetik parametre ise allellere ait frekans da ılımıdır. 31 genotipte incelenen 13 lokusa ait allel sayıları ve frekansları ekil 4.1 de verilmi tir. Buna göre VVS1 lokusunda 172 alleli, % ile en fazla frekansı vererek genotipler arasında en çok görülen allel olmu tur. Aynı ekilde 31 genotipte en fazla rastlanan allel, VVS3 lokusunda 212 (%40.32), VrZAG79 lokusunda 232 (% 22.58), VVMD5 lokusunda 230 (% 25.81), VVMD7 lokusunda 244 (% 22.58), VVMD27 lokusunda 194 (% 27.42), VVMD6 lokusunda 203 (% 27.42), VrZAG62 lokusunda 198 (% 27.42, VVMD31 lokusunda 216 (% 24.19), VrZAG67 lokusunda 133 (% 24.19), VVMD28 lokusunda 249 (%16.12), VVS2 lokusunda 146 (%16.13), allelleri olmu tur. Çalı ılan genotipler arasında VVS3 lokusundaki 212, VVS4 lokusundaki 172, VVMD5 lokusunda 230, VVMD6 lokusunda 203 allelleri, Türk çe itleri dahil Cabernet Sauvignon ve/veya Merlot çe itlerinde de görülmü tür. Bundan dolayı bunlara sadece Türk çe itlerine ait allellerdir ifadesi kullanılamaz. Ancak, VVS2 lokusundaki en fazla görülen 146, VVMD7 lokusunda 244, VVMD27 lokusunda 194, VVMD28 lokusunda 249, VVMD31 lokusunda 216, VrZAG62 lokusunda 198, VrZAG67 lokusunda 133, VrZAG79 lokusunda 232 allelleri sadece çalı mada Türk çe itlerinde görülen alleller olmu tur.
75 59 Allel frekansı 0,50 0,40 0,30 0,20 0,10 0,00 VVS Allel (bp) Allel frekansı 0,25 0,20 0,15 0,10 0,05 0,00 VVS Allel (bp) Allel frekansı 0,50 0,40 0,30 0,20 0,10 0,00 VVS Allel (bp)
76 60 Sekil 4.1. Çalı ılan lokuslarda tespit edilen allel frekanslarının da ılımı Allel frekansı 0,30 0,25 0,20 0,15 0,10 0,05 0,00 VVS Allel (bp Allel frekansı 0,30 0,25 0,20 0,15 0,10 0,05 0,00 VVMD Allel (bp) Allel frekansı 0,30 0,25 0,20 0,15 0,10 0,05 0,00 VVMD Allel (bp)
77 61 Sekil 4.1. (devamı) Allel frekansı 0,25 0,20 0,15 0,10 0,05 0,00 VVMD Allel (bp) Allel frekansı 0,30 0,25 0,20 0,15 0,10 0,05 0,00 VVMD Allel (bp) 0,20 VVMD28 Allel frekansı 0,15 0,10 0,05 0, Allel (bp)
78 62 Sekil 4.1. (devamı) 0,30 VVMD31 Allel frekansı 0,20 0,10 0, Allel (bp) Allel frekansı 0,30 0,25 0,20 0,15 0,10 0,05 0,00 VrZAG Allel (bp) Allel frekansı 0,30 0,25 0,20 0,15 0,10 0,05 0,00 VrZAG Allel (bp)
79 63 ekil 4.1. (devamı) Allel frekansı 0,25 0,20 0,15 0,10 0,05 0,00 VrZAG Allel (bp) ekil 4.1. (devamı) Genotiplere ait genetik benzerlik de erleri Çizelge 4.4 te verilmi tir. Buna göre en yüksek genetik benzerlik de eri T15 [17] ve Y3 [18], pek 13[14] ve pek 25 [15], Osmanca 26 [10] ve 40 [12] arasında (1.000) bulunmu tur. Bunu takiben Pembe Gemre 6 [21] ve 12 [23] (0.962), Pembe Gemre 6 [21] ve 11 [22] (0.923), Pembe Gemre 11 [22] ve 12 [23] (0.962), pek 4 [13] ve pek 13 [14] ile pek 4 [13] ve pek 25 [15] arasında (0.923), Sultani Çekirdeksiz S4 [19] ve S6 [20] (0.923), Osmanca 26 [10] ve 39 [11] ile 40 [12] ve 39 [11] arasında (0.885), Sultani Çekirdeksiz T3 [16] ve T15 [17] ile T3 [16] ve Y3 [18] arasında (0.885) gibi yüksek bir genetik benzerlik de eri gözlenmi tir Payla ılan allel oranı en az de eriyle Kozak Siyahı [2] ve Foça Karası (M) [9], Foça Karası (M) [9] ve Siyah Gemre [26], Foça Karası (M) [9] ve Gökçe Gemre [28], Kokulu Misket [3] ve Siyah Gemre
80 64 [26] ile Osmanca 39 [11] ve Sultani Gemre [27] arasında olmu tur. Aydın, Çanakkale ve zmir (Mordo an) illerinden alınan aynı isimli çe itler (homonim), farklı bulunmu tur. Buna göre farklı illerin aynı isimli çe itleri arasındaki genetik benzerlik de eri, Foça Karası (Aydın- Çanakkale) [7-8], Foça Karası (Aydın- zmir) [7-9] ve Foça Karası (Çanakkale- zmir) [8-9] ve Dumanlı Gemre [24-25] çe itlerinde sırasıyla 0.423; 0.423; 0.385, olarak bulunmu tur. Bu nedenle bu çe itlerin farklı oldu u sonucu ortaya çıkmaktadır. Yani bu çe itler homonim bulunmu tur. Ege Bölgesi asma genotiplerine ait dendogram ekil 4.2 de verilmi tir. Dendogram incelendi inde 2 ana grup göze çarpmaktadır. Genotiplerin büyük kısmını içeren birinci grup içinde çok sayıda alt grup gözlenmi tir. Di er grupta ise Osmanca çe idine ait 26 ve 40 no lu klonlarının payla ılan allel oranları ile de erlendirilen genetik benzerlikleri açıkça görülmektedir. Osmanca çe idine ait klonlar ile birlikte yerli araplık çe itlerimizden Foça Karası (09), Foça Karası (17), Foça Karası (M) çe itleri birlikte yer almı tır. Dendogramda Sultani Çekirdeksiz çe idine ait tiplerden T15 ve Y3 tiplerinin, pek üzüm çe idinin 13 ve 25 no lu klonların genetik benzerlikleri açıkça görülmektedir. Aynı ekilde Pembe Gemre çe idinin 6 ve 12 no lu klonları çok yakın genetik benzerlikte olduklarını söyleyebiliriz. Pembe Gemre çe idine ait 11 no lu klon da bu iki tipe genetik yakınlı ı nedeniyle dendogramda yakın ba lanmı tır. pek üzüm çe idinin 13 ve 25 no lu klonlarına ba lanan 4 no lu klonun da bu iki klona genetik benzerlikleri görülmektedir. Sultani Çekirdeksiz çe idine ait T15 ve Y3
81 65 tiplerine ba lanan T3 de bu iki tipe genetik yakınlı ı nedeniyle dendogramda yakın ba lanmı tır. Yine genetik olarak yakın bulunan S4 ve S6 tiplerinin de kendi arasında çok yakın genetik benzerlikte olduklarını söyleyebiliriz. Kurutmalık Özelli e sahip T3, T15, Y3 ile sofralık özelliklere sahip olan S4, S6 arasında arasında de i en benzerlik oranıyla genetik olarak birbirlerinden farklılık göstermi lerdir. Buca Razakısı ve Aydın Razakısı arasında gibi yakın bir genetik benzerlik de eri dendogramda da görülmektedir. Çalı madaki Gemre aksesyonları (Dumanlı Gemre (kol.), Halis Gemre, Gökçe Gemre ve Siyah Gemre, Dumanlı Gemre (os.), Sultani Gemre) aralarında genetik olarak benzerlik görülmemi ve dendogramda farklı alt gruplarda yer almı lardır.
82 Çizelge 4.4 Ege Bölgesine ait bazı önemli üzüm çeúitleri,tipleri ve klonlarının genetik benzerlik de erleri (Çeúitlerin numaralandırma sırası Çizelge 3.1 e göre yapılmıútır) 66
83 ekil 4.2. Ege Bölgesi asma genotiplerine ait dendogram 67
84 68 5. TARTI MA VE SONUÇ Bu çalı mada Manisa Ba cılık Ara tırma Enstitüsü nce klon seleksiyonu sonucu seçilmi Sultani Çekirdeksiz üzüm çe idine ait 5 tip (T3, T15, Y3, S4, S6 no lu), Pembe Gemre (6, 11, 12, no lu), Osmanca (26, 39, 40 no lu), pek üzüm çe itleri (4, 13, 25 no lu) ne ait 9 klon ve Ege Bölgesi için önemi olan 15 yerel çe it (Kozak Beyazı, Kozak Siyahı, Kokulu Misket, Buca Razakısı, Aydın Razakısı, Dumanlı Gemre (kol.) Dumanlı Germe (os.), Siyah Gemre, Sultani Gemre, Gökçe Gemre, Halis Gemre, Foça Karası (09) Foça Karası (17), Foça Karası (M), Bornova Misketi) ile 2 referans çe it (Cabernet Sauvignon ve Merlot) olmak üzere toplam 31 üzüm çe idinin (Vitis vinifera L.) DNA düzeyinde allel profilleriyle genetik tanımlanması yapılmı tır. Bu çalı mada kullanılan 16 mikrosatelit marköründen 13 adedi ile aralarındaki genetik benzerlik/farklılıklar belirlenmi tir (bunun için moleküler markörlerden mikrosatelitler kullanılmı tır). Mikrosatelitler; çe it tanımlaması, sinonim ve homonimlerin belirlenmesi gibi konularda birçok ara tırıcı tarafından kullanılmı ve kabul görmü tür (Dangl et al. 2001, Regner et al. 2001, Reale et al. 2002, This et al. 2004). Bu çalı mada genotiplerin moleküler analizi, SSR tekni i ile poliakrilamid jel kullanılarak yapılmı tır. Jel sonuçlarının gümü boyama ile görüntülenmesi ve sonuçların de erlendirilmesi bu ekilde gerçekle tirilmi tir. SSR la ortaya konulan allel büyüklüklerinin tespit edilmesinde gümü boyama, di er metotlar arasında güvenilir bir alternatiftir. Radyoaktif i aretli metodlara göre daha az tehlikeli ve daha
85 69 hızlıdır. Ayrıca otomatik florasan i aretlilere göre daha ucuzdur. Cipriani ve arkada ları (1994) e göre; amplifikasyon ürünleri, aynı zamanda yüksek çözünürlüklü agaroz jelde etidyum bromid ile görüntülenerek de analiz edilebilece ini ancak allel büyüklüklerinin güvenilir bir ekilde belirlenemeyece ini ve bu metot ile küçük farklılıkların akrilamid jelde oldu u gibi görülemeyece ini belirtmi lerdir (Bowers et al., 1996). Yukarıda adı geçen genotiplerin 13 SSR lokusu ile analiz edilmesi sonucu toplam allel sayısı 143 olarak belirlenmi tir. Allel sayıları lokuslarda 6 dan (VVS3) 15 e (VrZAG62) kadar de i mi ve ortalama 11 de erini vermi tir. Bu de erler, Bowers ve arkada larının 1999 yılında yaptıkları çalı mada, 51 üzüm çe idinin 12 mikrosatelit lokusta analizi sonucunda 4 ilâ 19 allel; Arroyo-Garcia ve Martinez- Zapater (2004), 41 çe itte 11 mikrosatelit lokusta 4 ilâ 12 allel; Fatahi ve arkada ları (2003), 62 genotip 9 mikrosatelit lokusta 4 (scu14vv) ilâ 16 (VVS2) (ortalama 11.4) allel; Vouillamoz ve arkada ları (2006), 116 aksesyon 12 mikrosatelit lokusta 6 (VVMD24) ile 16 (VVMD28 ve VVMD32) arasında de i en (ortalama 11.9) allel belirlemi lerdir. Çalı mamızdaki allel sayıları, belirtilen çalı malardaki ortalama allel sayılarıyla parelellik göstermektedir. Ara tırmada 16 primerin laboratuarda amplifikasyon standardizasyonu yapılarak örnekler üzerinde ara tırmalar yürütülmü tür. VVMD24 primerinde PCR reaksiyonlarındaki optimize çalı malarına ra men sonuç elde edilememi tir. VVMD17 primerinde ise null allel frekansı (r: 0.207) beklenen de erin üzerinde çıkarak null allel göstermi bu nedenle istatistik analizlere dahil edilmemi tir. Ayrıca VrZAG29 primeri de istatistik de erlendirmelerin dı ında bırakılmı tır.
86 70 Ara tırmada, beklenen heterozigotluk (He) ve gözlenen heterozigotluk (Ho) oranı ortalaması sırasıyla ve olarak bulunmu tur. VVS2, VVS3, VVS4 lokuslarında gözlenen heterozigotluk (Ho) yüksek bulunurken, VVS1, VVMD5, VVMD6, VVMD7, VVMD27, VVMD28, VVMD31, VrZAG62, VrZAG67, VrZAG79 lokuslarında ise beklenen heterozigotluk, (He) gözlenen heterozigotlu a (Ho) göre az bir farkla yüksek bulunmu tur. He de eri yüksek bulunan bu lokuslarda, null allellerin frekansı (r) pozitif olmakla birlikte de erlerinin dü ük olması nedeni ile null allel varlı ından üphelenilmemektedir (Çizelge 4.3). Tespit olasılı ı (PI) de eri hemen hemen tüm lokuslarda, Sefc et al. (2001) nın belirti i gibi 0.05 de erinden yüksek çıkmı tır. Bu da, bu seçilen mikrosatelit markörler asmada gerçekten yüksek derecede polimorfik olduklarını göstermektedir. En dü ük tespit olasılı ı (PI) de eri ile VVMD28 ve VrZAG79 (sırası ile PI de erleri ve ) markörleri çalı ılan mikrosatelit markörler arasında en fazla bilgi verici markörler iken PI de eri ve 6 allel sayısıyla VVS3 en az bilgi verici lokus olarak tespit edilmi tir. Yapılan di er çalı malara baktı ımızda, en fazla bilgi verici markörler VVS2, VVMD28 ( elli et al. 2007), VVMD14, VVMD28, VVMD36 (Crespan and Milani 2001), VVMD5, VVMD27, VVMD28, VVS2, VVS5 ve VrZAG67 (Ibáñez et al. 2003) olmu tur. Ara tırmada kullanılan 31 genotipte çalı ılan lokuslar de erlendirildi inde, VrZAG62 lokusu 15 ile en çok allel veren lokus olmu tur. Aynı lokus, Zulini et al. (2002), Karata et al. (2007) tarafından yaptıkları çalı malarda da en yüksek sayıda alleli vererek çalı mamızı
87 71 destekler nitelikte olmu tur. Bunu 14 allel ile VrZAG79 ve VVMD28 lokusları izlemi tir. VVMD28 in en fazla allel sayısına sahip lokus oldu unu Crespan ve Milani (2001), Vouillamoz ve arkada ları (2006) yaptıkları ara tırmada bildirmi lerdir. Reale ve arkada ları (2006), yaptıkları çalı mada en fazla alleli VrZAG79 lokusunun verdi ini ve en fazla allel sayısıyla da en çok bilgi veren lokuslar arasında yer aldı ını belirtmi tir. Bu sonuç çalı mamız sonucuna paralellik göstermektedir. En az allel veren lokus ise 6 allel ile, VVS3 lokusu olmu tur. Lokuslarda en fazla görülen allellere bakıldı ında, VVS1 lokusunda % allel frekansı ile 172 alleli, genotipler arasında en çok görülen allel olmu tur. Aynı ekilde 31 genotipte en fazla rastlanan allel, VVS3 lokusunda 212 (% 40.32), VrZAG79 lokusunda 232 (% 22.58), VVMD5 lokusunda 230 (% 29.03), VVMD7 lokusunda 244 (% 22.58), VVMD27 lokusunda 194 (% 27.42),VVMD6 lokusunda 203 (% 27.42), VrZAG62 lokusunda 198 (% 27.42), VVMD31 lokusunda 216 (% 24.19), VrZAG67 lokusunda 133 (% 24.19), VVMD28 lokusunda 249 (% 16.12), VVS2 lokusunda 146 (% 16.13) allelleri olmu tur. Çalı ılan genotipler arasında VVS3 lokusundaki 212, VVS4 lokusundaki 172, VVMD5 lokusunda 230, VVMD6 lokusunda 203 allelleri, Türk çe itleri dahil Cabernet Sauvignon ve/veya Merlot çe itlerinde de görülmü tür. Bundan dolayı bunlara sadece Türk çe itlerine ait allellerdir ifadesi kullanılamaz. Ancak, VVS2 lokusundaki en fazla görülen 146, VVMD7 lokusunda 244, VVMD27 lokusunda 194, VVMD28 lokusunda 249, VVMD31 lokusunda 216, VrZAG62 lokusunda 198, VrZAG67 lokusunda 133, VrZAG79 lokusunda 232 allelleri sadece çalı mada Türk çe itlerinde görülen alleller olmu tur. Bowers ve arkada larının 1996
88 72 yılında Kaliforniya daki koleksiyonda bulunan 77 üzüm çe idinde yapmı oldukları çalı mada, VVMD7 lokusundaki 245 allelinin sadece Yunanistan kökenli bir sofralık çe itte görüldü ünü bildirmi tir. VVMD7 lokusundaki 245 allelinin sadece Yunanistan çe itlerinde görülmedi i, çalı mamızda yer alan yerel çe itlerde de bulundu u yapılan bu çalı ma ile tespit edilmi tir. Yine aynı lokustaki 239 allelinin araplık çe itlerde sofralık çe itlerden daha yüksek bir allel frekansı gösterdi ini, buna kar ılık 249 allelinin de sofralık çe itlerde daha fazla görüldü ü belirtilmi tir. Çalı mamızda da aynı lokustaki 239 alleline, ço u araplık çe itte rastlanırken 24 sofralık çe itten sadece 1 inde görülmü tür. 249 alleli ise sadece sofralık çe itlerde görülmü tür. Ara tırmamızda da aynı lokustaki 239 ve 249 allellerindeki durum literatürde verilen sonuçlarla paralellik göstermi tir Çalı ılan genotiplerde analiz edilen 13 lokusta toplam 143 allel tespit edilmi tir. Allel büyüklüklerine bakıldı ında, Sultani Çekirdeksiz çe idinin T15 ve Y3 tipleri, pek çe idinin 13 ve 25, Osmanca üzüm çe idinin 26 ve 40 no lu klonları. her lokusta aynı allelleri verirken; Pembe Gemre 6 ve 12 no lu klonları sadece VrZAG62 lokusunda 1 allel; tarafından ayrılmı, di er 143 allelde aynı allel büyüklü ünü göstermi lerdir. Bu 1 allel farklılık, iki klondan birinde VrZAG62 lokusunda meydana gelen bir somatik mutasyonla ya da klonların poliklonal orijin olmasıyla açıklanabilir. Bütün bunlar moleküler genetik analiz çe itlerin klonal veya poliklonal özellikte olup olmadıklarını anlamamıza imkan verir (Vignani et al., 1996, Silvestroni et al., 1997). Bu yüzden Pembe Gemre nin 6 ve 12 no lu klonları ampelografik özellikler de göz önüne alınarak benzer genetik özellikte olduklarını
89 73 kabul edebiliriz; Sultani Çekirdeksiz çe idinin S4 ve S6 tipleri VVMD31 lokusunda 2 allel, Pembe Gemre 6, 11 ve 12 no lu klonları 2 allel, pek 4, 13 ve 25 no lu klonları 2 allel farklı, Sultani Çekirdeksiz çe idinin T3, T15 ve Y3 tipleri VVMD6 ve VVMD31 lokuslarında 3 allel; Osmanca 26, 39, 40 no lu klonları arasında 3 allel bakımından farklılık göstermi tir. Karaa aç (2006) tarafından yapılan çalı mada, Tekirda ve Gaziantep illerinden alınan Rumi çe itlerinde 3 allelde (VVMD31 ve VMC3B10 lokuslarında) farklılık görülmü tür. Bu farklılı ı dü ük bir ihtimal dahi olsa, bu lokuslardaki 3 ayrı yerde bir somatik mutasyonun gerçekle mi olabilece i eklinde açıklamı tır. Gerçekte bu ihtimalin olması, bu iki çe idin uzun yıllar boyunca aynı yerde yeti tirildikleri ve zaman içerisinde birden çok somatik mutasyona maruz kalmı olmaları anlamına geldi ini ve bu iki çe idin de aynı oldu unu belirtmi tir. Bu da, bu yerel çe itler için muhtemel bir olasılıktır. Bu yüzden Sultani Çekirdeksiz çe idinin S4 ve S6 tipleri, Pembe Gemre 6, 11 ve 12 no lu klonları, pek çe idinin 4, 13 ve 25 no lu klonları, Sultani Çekirdeksiz çe idinin T3, T15 ve Y3 tipleri; Osmanca 26, 39 ve 40 no lu klonlarının da benzer genetik özellikte olduklarını söyleyebiliriz. Buca Razakısı ve Aydın Razakısı arasında ise 5 allel farklı bulunmu tur. Buca Razakısı ve Aydın Razakısı incelenen 13 mikrosatelit lokusta, genetik benzerlik de erini verdi i görülmektedir. Bu nedenle bazı lokuslardaki allel farklılı ından dolayı farklı oldu u söylenebilir. elli ve arkada larının (2007), Anadolu dan alınan Dimrit ve Gemre üzüm gruplarından olu an çe itlerin mikrosatelit tanımlaması ve genetik farklılıklarını inceledikleri çalı mada, ortak olan Siyah Gemre (Gördes-Manisa) üzüm çe idi aynı lokuslarda kar ıla tırılmı tır ve
90 74 çalı mada ve bu tezde ortak çalı ılan toplam 7 lokustan, sadece.vvmd7 ve VVMD27 lokuslarında 2 allel aynı bulunmu tur. 7 mikrosatelit lokusta, % oranında allelleri payla tı ı görülmektedir. Yine çalı madaki Siyah Gemre (Konya) üzüm çe idi ortak çalı ılan lokuslarla kar ıla tırılmı VVS2, VVMD5, VVMD7, VVMD27 lokuslarında 6 allel aynı, di er lokuslarda farklı bulunmu tur. Her iki çalı mada ortak kullanılan 7 mikrosatelit lokusta % oranında allelleri payla tı ı görülmü tür. Bu nedenle tüm çe itlerin farklı oldu u görülmü tür. Gördes-Manisa, Konya orijinli Siyah Gemre çe idi ve bu çalı madaki zmir- Mordo an dan Manisa Ba cılık Ara tırma Enstitüsüne gelen Siyah Gemre üzüm çe itleri aynı isimli olmalarına ra men genetik olarak benzer bulunmamalarının nedeni bu çe itlerin Türkiye nin farklı bölgelerinden alınmı olmasından kaynaklanıyor olabilece i dü ünülmektedir. Çalı mamızda kullanılan referans çe itler Cabernet Sauvignon ve Merlot di er ara tırıcılar (Lin and Walker, 1998, Bowers et al. 1999, Fatahi et al. 2003, Karaa aç, 2006, elli et al. 2007) tarafından da çalı ılmı ve 15 lokusta da aynı lokustaki iki allel arasındaki fark çalı madaki sonuçlarla paralel bulunmu tur. Çe itler arasındaki ampelografik özellikler Çizelge 3.1 de verilmi tir. Buna göre, Dumanlı Gemre (kol.) ve Dumanlı Gemre (os.) çe itleri, farklı ara tırıcılar tarafından incelenen ampelografik özellikler bakımından aynı isimli bu çe itlerde tad, verimlilik ve salkım a ırlıkları bakımından farklı gruplarda yer almı lardır di er ampelografik özelliklerde uyu ma görülmü tür. Aydın, Çanakkale ve zmir (Mordo an) illerinden alınan Foça Karası çe itleri, Dilli (2008),
91 75 tarafından de erlendirilmi ve yine aralarında tane ekli, tane rengi ve verimlilik bakımından ayrılmı ve di er ampelografik özellikler örtü mü tür. Buca Razakısı ve Aydın Razakısı çe itleri, tane rengi, tane a ırlı ı, de erlendirme ekli, olgunluk zamanı özellikleri bakımından aynı, verimlilik, salkım a ırlı ı, tane ekli, tad özellikleri bakımından farklılık göstermi lerdir. Mikrosatelit markörlerle yapılan genetik analiz ile ampelografik çalı ma sonucu kar ıla tırıldı ında ise, Dumanlı Gemre ve Foça Karası çe itleri ampelografik ve genetik analiz sonuçları bu iki örnekte birbiriyle uyumlu olmu tur. Her iki sonuçta da farklı oldukları görülmü tür. Bu nedenle bu çe itlerin farklı oldu u sonucu ortaya çıkmaktadır. Yani bu çe itler homonim bulunmu tur. Ara tırmada elde edilen sonuçların kısa özeti: - Osmanca 26 ve 40, pek 13 ve 25 no lu klonları ve Sultani Çekirdeksiz çe idine ait T15 ve Y3 tipleri ve aynı çe ide ait klonlar kendi aralarında aynı bulunmu tur. - Sultani Çekirdeksiz çe idine ait tiplerden T3, T15, Y3 (Kurutmalık özelli e sahip) ile S4, S6 (sofralık özelli i belirgin) isimli genotipler kar ıla tırıldı ında arasında de i en genetik benzerlik de erleri sonucu birbirlerinden farklılık göstermi lerdir - Ara tırmada yer alan Gemre aksesyonları (Dumanlı Gemre (kol.), Halis Gemre, Gökçe Gemre, Siyah Gemre, Dumanlı Gemre (os.) ve Sultani Gemre) arasında genetik olarak benzerlik görülmemi tir. - Ara tırmadaki Aydın, Çanakkale ve zmir (Mordo an) illerinden alınan aynı isimle alınmı Foça Karası çe itleri arasında ve Manisa Ba cılık Ara tırma Enstitüsü nde ayrı parsellerde bulunan Dumanlı
92 76 Gemre üzüm çe itlerinin de farklı oldukları görülmü tür ve homonim duruma rastlanmı tır. Bu çe itler ampelografik ve genetik olarak tamamen farklı bulunmu tur. - Farklı yıllarda ve farklı ara tırıcılar tarafından yapılan ampelografik çalı ma sonuçları, çalı mamızdaki genetik analiz sonuçları büyük bir ço unlukla uyum içerisindedir. - Mikrosatelit markörlerin, çalı mamızdaki genotiplerin genetik tanımlamalarında uygun oldu u belirlenmi tir. Bunlardan VVMD28 ve VrZAG79 markörleri, çalı ılan mikrosatelit markörleri arasında en fazla bilgi verici markörlerdir. - Ara tırmada Avrupa Birli i Genres CT96 No81 projesi çerçevesince geli tirilen ve OIV tarafından da onaylanan asma çe it analizi için minimum standart mikrosatelit markör seti olarak kabul edilen markörleri de kapsayacak ekilde 13 markörle 31 yerel üzüm çe it, tip ve klonların kimlik tespitleri gerçekle tirilmi tir. Ülkemizde de hem yerel hem de ticari öneme sahip uluslararası üzüm çe itlerinin SSR veri bankası olu turulmalıdır. Böylece bu çalı ma ve benzer çalı malardan elde edilen sonuçlar ulusal veri bankasında de erlendirilebilecektir. Bu veri bankasından üzüm ve üzüm ürünleri ticaretinden, fidancılık alanına kadar pek çok sektörde ismine do rulu un belirlenmesinde yararlanılabilecektir. Yine ıslah çalı malarından, üzüm çe it ve klonlarının tescil ve sertifikasyon a amalarına kadar bir çok alanda veri bankası kullanılabilecektir.
93 77 KAYNAKLAR D Z N A ao lu, Y.S., 1999, Bilimsel ve Uygulamalı Ba cılık (Asma Biyolojisi). Kavaklıdere E itim Yayınları, No: s, Ankara. A ao lu, Y.S. ve Ergül, A., 1999, Amasya Tipi Üzüm Çe itlerinin RAPD (Random Amplified Polymorphic DNA) Tekni i ile Genetik tanımlamaları. Türkiye III. Ulusal Bahçe Bitkileri Kongresi Eylül 1999, Ankara, s: A ao lu, Y.S., Söylemezo lu G., Çalı kan, M. ve Ergül, A. 1999, Türkiye de Yeti tirilen Razakı Üzüm Çe idi Ekotiplerinin Elektroforetik Tanımlamaları Üzerinde Ara tırmalar. Türkiye III. Ulusal Bahçe Bitkileri Kongresi Eylül, s: Ankara. Akkak, A., Boccacci, P. and Botto, R., 2007, Cardinal Grape Parentage: A Case Of A Breeding Mistake, Genome 50: Alleweldt G., 1988, The Genetic Resources Of Vitis. Genetic And Geographic Origin of Grape, Their Prime Names and Synonims. 2nd ed. Federal Research Center for Grapewine Breeding, Geilweilerhof, Germany. Anonim, 1997, Standart Üzüm Çe itleri Katalo u, Tarım ve Köyi leri Bakanlı ı, Aradhya, M.K., Dangl, G.S., Prins, B.H., Boursiquot, J.M., Walker, A.M., Meredith, C.P., Simon, C.J., 2003, Genetic Structure and Differentiation in Cultivated Grapes Vitis Vinifera L. Genet. Res. Camb. 81: Plant & Animal Genomes VIII. Conference, Town & Country Hotel, San Diego, CA.
94 78 KAYNAKLAR D Z N (devam) Arroyo-Garcia, R., L. Ruiz-Garcia, L. Boulling, R. Ocete, M. A. López, C. Arnold, A. Ergul, G. Söylemezo lu, H.. Uzun, F. Cabello, J. Ibáñez, M. K. Aradhya, A. Atanassov, I. Atanassov, S. Balint, J. L. Cenis, L. Costantini, S. Gorislavets, M. S. Grando, B. Y. Klein, P. McGovern, D. Merdinoglu, I. Pejic, F. Pelsy, N. Primikirios, V. Risovannaya, K. A. Roubelakis-Angelakis, H. Snouss, P. Sotiri, S. Tamhankar, P. This, L. Troshin, J. M. Malpica, F. Lefort, and J. M. Martinez-Zapater., 2006, Genetic Evidence for The Existence of ndependent Domestication Events n Grapevine. Molecular Ecology, 15(12), Balerias-Couto M.M. and Eiras-Dias J.E., 2006, Detection and Identification of Grape Varieties in Must and Wine Using Nuclear and Chloroplast Micosatellite Markers, Analytica Chimica Acta. Vol. 563, Issues 1-2, pp , March Bowers, J.E., Dangl, G.S., Vignani, R., Meredith, C.P., 1996, Isolation and Characterization of New Polymorphic Simple Sequence Repeat Loci in Grape (Vitis vinifera L.). Genome, 39: Bowers, J.E.ve Meredith C.P., 1997, The Parantage of Classic Winw Grape: Cabernet Sauvignon. Nat. Genet. 16: Bowers J.E., Dangl, G.S., Meredith, C.P., 1999a, Development and Characterization of Additional Microsatellite DNA Markers for Grape. Am J Enol Vitic. 50: Bowers J.E., Boursiquat J.M., This, P., Chu, K., Johnsson, H., Meredith, C.P., 1999b, Historical Genetics: The Parantage of Chardonnay, Gamay, and other Wine Grapes of Northeastern of France. Science, Vol 285:
95 79 KAYNAKLAR D Z N (devam) Boz, Y.ve Çelik, S., 1998, Melezlemeyle Elde Edilen Sofralık Ümitvar Bazı Üzüm Çe it ve Adayları ile Bunların Ebeveyn Çe itlerinin Ampelografik Özelliklerinin Belirlenmesi. 4. Ba cılık Sempozyumu Bildirileri s. Yalova. Brookfield, J.F.Y., 1996, A Simple New Method for Estimating Null Allele Frequency from Heterozygote Deficiency. Mol Ecol 5; Cabezas, J.A., Cervera, M.T., Arroyo-Garcia, R., Ibáñez, J., Rodrígues-Torres, I., Borrego, J., Cabello, F. and Martínez- Zapater, J.M., 2003, Garnacha and Garnacha Tintorera: Genetic Relationships and The Origin of Teinturier Varieties Cultivated in Spain. Am. J. Enol. Vitic., 54 (4), Collins GG. And Symons, R.H., 1999, Polymorphism in Grapevine DNA Detected by The RAPD-PCR Tecnique. Plant Mol Biol Rep. 11: Crespan, M. and Milani, N., 2001, The Muscats: A Molecular Analysis of Synonyms, Homonyms and Genetic Relationships within A Large Family of Grapevine Cultivars. Vitis, 41(1), Crespan, M., Cancellier, S., Costacurta, A., Guist, M., Carraro, R., Stefano, R. and Santangelo, S., 2003, Contribution to The Clearing Up of Synonymies in Some Groups of Italian Grapevine Cultivars. Proc. VIIIth IC on Grape, Acta Horticulturae, 603, Çelik, H., 2006, Üzüm Çe it Katalo u (Grape Cultivar Catalog). SUNF DAN A.. Mesleki Kitaplar Serisi : 3, 165 s. Ankara.
96 80 KAYNAKLAR D Z N (devam) Çelik, S., 1998, Ba cılık (Ampeloloji). Trakya Üniversitesi Tekirda Ziraat Fakültesi Bahçe Bitkileri Bölümü. Tekirda. Dangl, G.S., Mendum, M.L., Prins, B.H., Walker, A.M., Meredith, C.P. and Simon C.J., 2001, Simple Sequence Repeat Analysis of A Clonally Propagated Species: A Tool for Managing A Grape Germplasm Collection. Genome 44; Dice, L.R., 1945, Measures of The Amount of Ecological Association Between Species. Ecology, 26: v.26, p Dilli, Y., Kader, S., Yöresel Gemre Çe itleriyle Pembe Gemre Klonlarının Ampelografik Nitelikleri le Verim ve Kalite Özelliklerinin Kar ıla tırılması Üzerinde Bir Ara tırma. Türkiye 6. Ba cılık Sempozyumu Bildirileri. TEK RDA. Dilli, Y., 2008, Ege Bölgesindeki Bazı Önemli Üzüm Çe itleri, Tipleri ve Klonlarının Mikrosatelit (SSR) Markörleriyle Karakterizasyonu. (Sonuç Raporu). Manisa Ba cılık Ara tırma Enst. Müd. Basılmamı, Manisa. Doligez, A., Adam-Blondon, A.F., Cipriani, G., Digaspera, G., Louco, V., Merdino lu, D., Meredith, C.P., Riaz, S., Roux, C., This, P., 2006, An Integrated SSR Map of Grapevine Based on Five Mapping Sample. Theor Appl Genet. 113: Ecevit F. E.ve Kelen, M., Isparta (Atabey) de Yeti tirilen Üzüm Çe itlerinin Ampelografik Özelliklerinin Belirlenmesi Üzerine Bir Ara tırma. Tr. J. of Agriculture And Forestry
97 81 KAYNAKLAR D Z N (devam) Ergül, A. ve A ao lu, Y.S., 2001, Ülkemizde Asma Fidanı Üretiminde Kullanılan Bazı Amerikan Asma Anaçlarının Moleküler Benzerlik Analizi. A.Ü.Z.F. Tarım Bilimleri Dergisi. 7(2), Ankara. Dokuzo uz, M., 1964, Bahçe Bitkilerinin Islahında Klon Seleksiyonu. E. Ü. Z. F. Yayınları. 87. Ege Üniv. Matbaası. zmir. Ergül, A., Aras, E. S., Söylemezo lu, G. ve A ao lu, Y. S., 2002, Kalecik Karası Üzüm Çe idi Klonlarında AFLP (Amplified Fragment Length Polymorphism) Tekni i ile Polimorfizmin Belirlenmesi. Türkiye V. Ba cılık ve arapçılık Sempozyumu. 5-9 Ekim 2002, Bildiriler : Nev ehir. Faria, M.A., Magalhaes, R., Ferreira, M.A., Meredith, C.P. and Monteiro, F.F., 2000, Vitis Vinifera Must Varietal Authentication Using Microsatellite DNA Analysis (SSR). J Agric Food Chem., 48(4), Fatahi, R., Ebadi, A., Bassil, N., Mehlenbacher, S.A. and Zamani, Z., 2003, Characterization of Iranian Grapevine Cultivars Using Microsatellite Markers. Vitis, 42(4), Fidan, Y., Eri, A., Tamer, M. S., 1972, Güdül lçesi Ba cılı ı Geli me mkanları ve Önemli Üzüm Çe itlerinin Ampelografik Vasıfları Üzerinde Bir Ara tırma. Ankara Üniversitesi Ziraat Fakültesi Yıllı ı, 21: , Ankara.
98 82 KAYNAKLAR D Z N (devam) Fossati, T., Labra M., Castiglione, S., Failla, O., Scienza, A.and Sala, F., 2001, The Use of AFLP and SSR Molecular Markers to Decipher Homonyms and Synonyms in Grapevine Cultivars: The Case of The Varietal Group Known As Schiave. Theor Appl Genet., 102, Goto-Yamamoto, N., Mouri, H., Azumi, M. and Edwards, K.J., 2006, Development of Grape Microsatellite Markers and Mikrosatellite Analysis Including Oriental Cultivars. Am. J. Enol. Vitic., 57(1), Ibáñez, J., Andrés, M.T., Molino, A. and Borrego, J., 2003, Genetic Study of Key Spanish Grapevine Varieties Using Microsatellite Analysis. Am. J. Enol. Vitic. 54 (1); çi, B., 2007, Asma (Vitis Vinifera L.) da Genom Haritalaması: Önemli Morfolojik Karakterlere ve Fungal Kökenli Hastalıklara Yönelik AFLP ve SSR Linkage Gruplarının Olu turulması, Doktora Tezi, E.Ü. Fen Bilimleri Enstitüsü, 211s (yayınlanmamı ). Ilgın, C., Öztürk H., Kader S., Erdem A., Gökçay, E.,1999, Ege Bölgesinde Yeti tiricili i Yapılan Bazı Çekirdeksiz Üzüm Çe itlerine ait Tiplerin Belirlenmesi Üzerine Ara tırmalar Manisa Ba cılık Ara tırma Enst. Müd. Yayın no: 80. Kader, S., 2005, Osmanca Üzüm Çe idindeki Klon Seleksiyonu Çalı ması Sonucunda Seçilen Klonlar le Osmanca Çe idinin Ampelografik Özelliklerinin Kar ıla tırılması. Türkiye 6. Ba cılık Sempozyumu Bildirileri. Cilt: s. Tekirda.
99 83 KAYNAKLAR D Z N (devam) Kader, S.ve Öztürk, H., 2005, Razakı Üzüm Çe idinde Klon Seleksiyonu Çalı ması Sonucunda Seçilen Klonların Ampelografik Özellikleri ile Göz Verimliliklerinin Belirlenmesi. Türkiye 6. Ba cılık Sempozyumu Bildirileri. Tekirda. Kafkas, S., 2006, Adana Biyoteknoloji Günleri Sunu, Ç. Ü. Bahçe Bitkileri Bölümü, Adana. Kaplan, N., 1995, Diyarbakır ve Mardin llerinde Yeti tirilen Üzüm Çe itlerinin Ampelografik Özelliklerinin Saptanması Üzerine Bir Ara tırma. II. Bahçe Bitkileri Kongresi s. Adana. Karaa aç, E., 2006, Gaziantep li Asma Gen Potansiyelinin SSR (Simple Sequence Repeats) Markörlerle Moleküler Analizi. Ankara Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Anabilim Dalı. Doktora Tezi. 89s. (yayınlanmamı ). Karata, H., De irmenci D., Velasco R., Vezzulli S., Bodur Ç., A ao lu, Y. S., 2007, Microsatellite Fingerprinting of Homonymous Grapevine (Vitis Vinifera L.) Varieties in Neighboring Regions of South-East Turkey. Scientia Horticulturae 114: Kısakürek, H., 1950, Güney-Do u Anadolu ve Bilhassa Gaziantep Ba cılı ı ve Bu Bölgede Yeti en Ba lıca Üzüm Çe itlerinin Morfolojik Vasıfları ve ktisadi Önlemleri Üzerinde Ara tırmalar. Ankara Üniversitesi Basımevi. Lamboy, W.F., and Alpha, C.G. 1998, Using Simple Sequence Repeats (Ssrs) for DNA Fingerprinting Germplasm Accessions of Grape (Vitis L.) Species. J. Am. Soc. Hortic. Sci. 123(2):
100 84 KAYNAKLAR D Z N (devam) Lander, E.S., Green, P., Abrahamson, J., Barlow, Daly, M.J., Lincoln, S.E. and Newberg, L., 1987, MAPMAKER, An Interactive Computer Package for Constructing Primary Genetic Linkage Maps of Experimental and Natural Populations. Genomics, 1: Lin, H. and Walker, M.A., 1998, Identifying Grape Rootstocks with Simple Sequence Repeat (SSR) DNA Markers. Am. J. Enol. Vitic. 49(4), Lefort, F., Lall, M., Thompson, D. and Douglas G.C., 1998, Morphological Traits, Microsatellite Fingerprinting and Genetic Relatedness of a Stand of Elite Oaks ( Q. robur L.) at Tullynally, Ireland. Silvae Genetica, 47: 5-6. Lopes, M.S., Sefc K.M., Eiras-Dias, E., Steinkellner, H., Lamier da Camara Machado, M., Dacamara Machando, A., 1999, The Use of Microsatellites for Germplasm Management in Portuguese Grapevine Collection Theor Appl Genet. 99: Malossini, U., Grando, M.S., Roncador, I. and Mattivi, F., 2000, Parentage Analysis and Characterization of Some Italian Vitis Vinifera Crosses. Proc. VIIth IC on Grapevine Genetics and Breeding, Acta Horticulturae. No: 528; Manzo P. and Tamponi, G., 1979, Monografia di Cultivar di uve da Tavola. Istituto Sperimentale per la Frutticoltura. Roma.
101 85 KAYNAKLAR D Z N (devam) Marasalı, B., 1995, Üzüm çe itlerinin RFLP (Restriction Fragment Length Polymorphism) Tekni i ile Moleküler Tanımlaması. Biyoteknoloji ve Bitki Islahı Workshop Nisan, Gebze/Kocaeli, s: Mauro M.C., Strefler M., Weeden NF, Riech BI., 1992, Genetic Analysis of Restriction Fragment Length Polymorphisms in Vitis. J. Hered. 63: Merdinoglu, D., Butterlin, G., Baur, C., Balthazard, J., Bouquet, A. and Boursiquot, J.M., 2000, Comparison of RAPD, AFLP and SSR (microsatellite) Markers for Genetic Diversity Analysis in Vitis vinifera L.. Acta Horticulturae, 528, Minch, E., Ruiz-Linares, A., Goldstein, D.B., Feldman, M. and Cavalli- Sforza, L.L.,1995, Microsat (version 1.4d): A Computer Program for Calculating Various Statistics on Microsatellite Allele Data. Stanford. California, University of Stanford. Morgante, M., and Oliveri, A. M. 1993, PCR Amplified Microsatellites as Markers in Plant Genetics. Plant J. 3: Najafi, J., Alipanah, L., Ghareyazie, B., Mohmammadi, S.A:, Nazari, A.H., This, P., 2006, Genetic Diversity of Iranian and Sone of European Grapes Revealed by Microsatellite Markers. Iranian Jaurnal of Biotechnology, Vol. 4, No: 1, Nei, M., 1987, Molecular Evolutionary Genetics, Columbia University ress, New York.
102 86 KAYNAKLAR D Z N (devam) Oraman, M. N., 1941, Çavu Üzümünün Vatanı, Ampelografisi ve Biyolojisi Üzerinde Bir Ara tırma. Yük. Ziraat Enstitüsü Çalı malarından Sayı: 114. Ankara. Oraman, M.N., 1970, Ba cılık Tekni i, Ankara Üniversitesi Ziraat Fakültesi Yayınları, 415, Ankara, 283s. Özcan, S., Gürel, E., Babao lu, M., 2001, Bitki Biyoteknolojisi (Genetik Mühendisli i ve Uygulamaları). S.Ü. Vakfı Yayınları, 456s. Paetkau D., Calvert W., Stirling I. and Strobeck C., 1995, Microsatellite Analysis of Population Structure in Canadian Polar Bears. Mol. Ecol. 4; Parfitt D.E. and Arulsekar S., 1989, Inheritance and Isoenzyme Diversity for GPI and PGM Among Grape Cultivars. J. Amer. Soc. Hort. Sci. 114: Perold, A. J., 1927, A Treatise on Viticulture. London. Powell, W., Machray GG, Provan, J., 1996, Polymorphism Revealed by Simple Sequence Repeats. Trends in Plant Sci. 1: Regner, F., Wiedeck, E. And Stadlbauer, A., 2000, Differentiation and Identification of White Riesling Clones by Genetic Markers. Vitis 39(3), Regner, F., Stadlbauer, A., Eisenheld, C., 2001, Molecular Markers for Genotyping Grapevine and for Identifying Clones of Traditional Varieties. Acta Hort. 546,
103 87 KAYNAKLAR D Z N (devam) Regner, F., Hack, R. ve Santiago, L., 2006, Highly Variable Vitis Microsatellite Loci for The Identification of Pinot Noir Clones, Vitis, 45(2): Reale, S., Pilla, F. and Angiolillo, A., 2006, Genetic Analysis of The Italian Vitis Vinifera Cultivar Tintilia and Releated Cultivars Using SSR Markers. Journal of Horticultural Science & Biotechnology 81(6) Riaz, S., and Meredith, C.P, 2000, A Microsatellite Marker Based Linkage Map of Vitis vinifera. In Abstract of the International Plant and Animal Genome Conference VIII, held 9-12 January 2000, San Diego, Calif. Scherago International Inc, New York (Abstr. P. 132). Gizella, J., János, K., János, M., Anita, L.,. Júlia G. M., 2007, Analysis of Grapevine (Vitis vinifera L.) Varieties by Molecular Markers. XXX th OIV World Congress, June 2007, Budapest. Riaz, S., Garrison, K.E., Dangl, G.S. and Meredith, C.P., 2001, Mikrosatellite Markers for The Differentiation of Clones of Ancient Grape Cultivars. Plant & Animal Genomes IX Conference, Town & Country Hotel, San Diego, CA. Samancı, H., ve Uslu,., 1992, Türkiye de Yeti tirilen Razakı Çe it ve Tiplerinin Ampelografik Özellikleri Üzerinde Ara tırmalar. (Sonuç Raporu). Atatürk Bahçe Kültürleri Merkez Ara tırma Enstitüsü,. Yalova, 26 s.
104 88 KAYNAKLAR D Z N (devam) Sefc K.M., Regner F., Glossl J. and Steinkellner, H., 1998a, Genotyping of Grapevine and Rootstock Cultivars Using Microsatellite Markers. Vitis, 37(1), Sefc, K.M., Guggenberger, S., Regner, F., Lexer, C., Glössl, J. And Steinkellner, H., 1998b, Genetic Analysis of Grape Berries and Raisins Using Microsatellite Markers. Vitis, 37(3), Sefc, K.M., Lefort, F., Grando, M.S., Scott, K.D., Steinkellner, H. and Thomas, M.R., 2001, Microsatellite Markers for Grapevine: A State of The Art. In Molecular Biology and Biotechnology of Grapevine. Roubelakis-Angelakis K.A. ed., 1-29, Kluwer Academic Publishers. The Netherlands. Sefc, K.M., Lopes, M.S., Lefort, F., Botta, R., Robelakis-Angelekis, K.A., Ibanez, J., Pejic, I., Wagner, H.W., Glossl, J., Steinkller, H., 2000, Microsatellite Variability in Grapevine Cultuvars from Different European Regions and Evolution of Assignment Testing To Assess The Geographical Origin of Cultivars. Theor Appl Genet. 100: Sefc K.M., Renger F., Turetschek E., Glossl J., Steinkellner, H., 1999, Identification of Microsatellite Sequnce in Vitis Riparia and Their Applicability for Genotyping of Different Vitis Species. Genome, 42: Sefc K.M., Steinkellner, H., Wagner, H.W., Glossi, J., Regner, F., 1997, Application of Microsatellite Markers for Parantage Studies in Grapevine. Vitis 36:
105 89 KAYNAKLAR D Z N (devam) elli F., M. Bakır, G. Inan, H. Aygün, Y. Boz, A. S. Ya asın, C. Özer, B. Akman, G. Söylemezo lu, K. Kazan, A. Ergül, Simple Sequence Repeat-Based Assesment of Genetic Diversity in Dimrit and Gemre Grapevine Accessions from Turkey. Vitis, 46(4): Silvestroni, O., Dipietro, D., Interieri, C., Vignani, R., Fillipetti, I., Del Casino, C., Scal, M., Cresti, M., 1997, Defection of Genetic Diversity Among Clones of cv. Fortana (Vitis vinifera L.) by Microsatellite DNA Polimorphism Analysis. Vitis, 36: Söylemezo lu, G., A ao lu, Y.S., Marasalı, B., Ergül, A., Çalı kan, M., Türkben, C., 1998, Üzüm Çe itlerinin Yaprak Kökenli Kate ol Oksidaz (CO), Peroksidaz (PER) ve Esteraz (EST) zoenzimlerinden Yararlanılarak Tanımlanmaları. 4. Ba cılık Sempozyumu Bildirileri, Ekim s: Yalova. This, P., Jung, A., Boccacci, P., Borrego, J., Botta, R., Costantini, L., Crespan, M., Dangl, G.S., Eisenheld, C., Ferreira Monteiro, F., Grando, M.S., Ibanez, J., Lacombe, T., Laucou, V., Magalhaes, N., Meredith, C.P., Milani, N., Peterlunger, E., Regner, F., Zulini, L. and Maul, E., 2004, Development of a Standard Set of Microsatellite Reference Alleles for Identification of Grape Cultivars. Theoretical and Applied Genetics, 109, Thomas M.R. and Scott NS., 1993, Microsatellite Repeats in Grapevine Reveal DNA Polymorphism When Analyzed as Sequence- Tagged Sites. Theor Appl Genet. 68:
106 90 KAYNAKLAR D Z N (devam) Thomas, M.R., Cain, P. and Scott, N.S., 1994, DNA Typing of Grapevines: A Universal Methodology and Database for Describing Cultivars and Evaluating Genetic Relatedness. Plant Mol. Biol. 25; Ulanovsky, S., Gogorecna, Y., Martinez Detoda, F., Ortiz, J.M., 2002, Use of Molecular Markers Indefection of Synonymies and Homonymies in Grapevine (Vitis vinifera L.). Sci Hortic. 92: Uzun, H.. ve lter, E., 1993, Bazı Üzüm Çe itlerinin Yapraklarındaki Peroksidaz ve Kate ol Oksidaz zoenzimlerinden Te hisi Üzerinde Ara tırmalar. E.Ü. Ziraat Fakültesi Dergisi, Cilt 30, (3); Viala, P. et Vermorel, V., 1910, Traite General de Viticulture Ampelographie, Paris. Vignani, R., Scali, M., Masi, E. and Cresti, M. 2002, Genomic Variability in Vitis Vinifera L. Sangiovese Assessed by Microsatellite and Non-Radioactive AFLP Test. Electronic Journal of Biotechnology., Vol. 5 No: 1 (www. ejb.ucv.cl). Vouillamoz, J.F., McGovern, P.E., Ergul, A., Söylemezo lu, G., Tevzadze, G., Meredith, C.P. and Grando, M.S., 2006, Genetic Characterization and Relationships of Traditional Grape Cultivars from Transcaucasia and Anatolia. Plant Genetic Resources: Characterization & Utilization, 4(2), Wagner, H.W. and Sefc, K.M. 1999, IDENTITY 1.0. Centre for Applied Genetics, University of Agricultural Science, Vienna.
107 91 KAYNAKLAR D Z N (devam) Weeden, N.F., B.I. Reisch, M.H.F. Martens, 1988, Genetic Analysis of Isozymes Polymorphism in Grape. J. Amer. Soc. Hort. Sci.,113(5) Weising, K., Winter, P., Huttel, B., Kahl, G., 1998, Microsatellite Markers for Molecular Breeding. J. Crop. Prod. 1: Welsh, J., Ralph D., McClelland, M. 1995, DNA and RNA Fingerprinting Using Arbitrarily Primed PCR. In: Innis MA, Gelfand DH, Sninsky JJ, (eds), PCR Strategies, Academic Press, San Diego, pp Wolfe, W. H., 1976, Identification of Some Grape Varieties by Isozyme Banding Patterns. Amer. J. Enol. Vitic., 27(2): Ye,G.N., Söylemezo lu, G., Weeden, N.F., Lambow, W.F., Pool, R.M. and Reisch, B.I., 1998, Analysis of The Relationship Between Grape Cultivars, Sports and Clones via DNA Fingerprinting. Vitis 37(1), Zulini, L., Russo, L. and Peterlunger, E., 2002, Genotyping Wine and Table Grape Cultivars from Apulia (Suthern Italy) Using Microsatellite Markers. Vitis 41(4),
 Foça Karası")
 Dumanlı Ge. (os.")

108 92 EKLER EK 1. Jel görütülerinde yer alan genotiplerin isimleri C. Sauvignon Merlot Kozak Beyazı Kozak Siyahı Kokulu Misket Bornova Misketi Buca Razakısı Aydın Razakısı Foça Karası (09) Foça Karası (17) Foça Karası (M) Osmanca (26) Osmanca (39) Osmanca (40) pek (4) pek (13) pek (25) Sultani Çek. T3 Sultani Çek. T15 Sultani Çek. Y3 Sultani Çek. S4 Sultani Çek. S6 Pembe Gemre 6 Pembe Gemre 11 Pembe Gemre 12 Dumanlı Ge. (kol.) Dumanlı Ge. (os.) Siyah Gemre Sultani Gemre Gökçe Gemre Halis Gemre C M EK 2. Ara tırmada kullanılan bazı SSR primerlerine ait poliakrilamid jel görüntüleri. VVS1 Primerine ait poliakrilamid jel görüntüsü VVS2 Primerine ait poliakrilamid jel görüntüsü VVS3 Primerine ait poliakrilamid jel görüntüsü


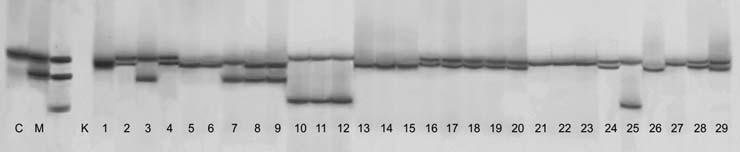

109 93 VVS4 Primerine ait poliakrilamid jel görüntüsü VVMD5 Primerine ait poliakrilamid jel görüntüsü VVMD6 Primerine ait poliakrilamid jel görüntüsü VVMD7 Primerine ait poliakrilamid jel görüntüsü


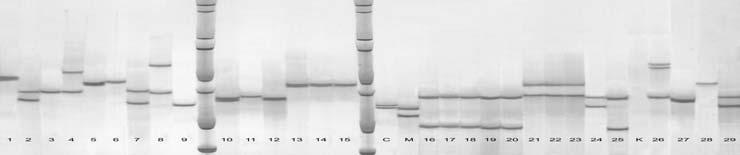

110 94 VVMD17 Primerine ait poliakrilamid jel görüntüsü VVMD27 Primerine ait poliakrilamid jel görüntüsü VVMD28 Primerine ait poliakrilamid jel görüntüsü VVMD31 Primerine ait poliakrilamid jel görüntüsü


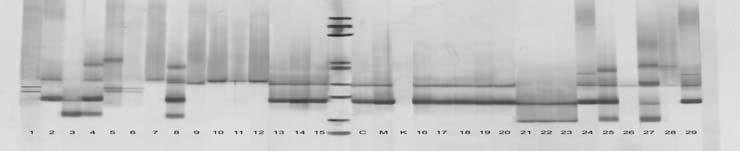

111 95 VrZAG29 Primerine ait poliakrilamid jel görüntüsü VrZAG62 Primerine ait poliakrilamid jel görüntüsü VrZAG67 Primerine ait poliakrilamid jel görüntüsü VrZAG79 Primerine ait poliakrilamid jel görüntüsü
ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ
 ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ KARA (SİYAH) ÜZÜM GRUPLARININ SSR(Simple Sequence Repeat) MARKÖRLERE DAYALI KARAKTERİZASYONU VE ÜLKE ASMA KAYNAKLARI İLE GENETİK İLİŞKİSİ
ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ KARA (SİYAH) ÜZÜM GRUPLARININ SSR(Simple Sequence Repeat) MARKÖRLERE DAYALI KARAKTERİZASYONU VE ÜLKE ASMA KAYNAKLARI İLE GENETİK İLİŞKİSİ
ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ
 ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ GAZİANTEP İLİ ASMA GEN POTANSİYELİNİN SSR (SIMPLE SEQUENCE REPEATS) MARKÖRLERLE MOLEKÜLER ANALİZİ EDA KARAAĞAÇ BAHÇE BİTKİLERİ ANABİLİM DALI ANKARA
ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ GAZİANTEP İLİ ASMA GEN POTANSİYELİNİN SSR (SIMPLE SEQUENCE REPEATS) MARKÖRLERLE MOLEKÜLER ANALİZİ EDA KARAAĞAÇ BAHÇE BİTKİLERİ ANABİLİM DALI ANKARA
TEZ ONAYI Funda YILDIRIM tarafından hazırlanan Ankara ve Çankırı İlleri Asma Gen Kaynaklarının SSRs (Simple Sequence Repeats) a Dayalı Genetik Karakte
 a Dayalı Genetik Karakte") ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ ANKARA ve ÇANKIRI İLLERİ ASMA GEN KAYNAKLARININ SSRs (SIMPLE SEQUENCE REPEATS) a DAYALI GENETİK KARAKTERİZASYONU Funda YILDIRIM BAHÇE BİTKİLERİ
ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ ANKARA ve ÇANKIRI İLLERİ ASMA GEN KAYNAKLARININ SSRs (SIMPLE SEQUENCE REPEATS) a DAYALI GENETİK KARAKTERİZASYONU Funda YILDIRIM BAHÇE BİTKİLERİ
ÖZEL EGE L SES. HAZIRLAYAN Ö RENC LER: Tayanç HASANZADE Ahmet Rasim KARSLIO LU. DANI MAN Ö RETMEN: Mesut ESEN Dr. ule GÜRKAN
 ÖZEL EGE L SES BAZI ISKARTA BALIKLARIN (Isparoz, Hani) ETLER NDEN ALTERNAT F GÜBRE YAPIMI VE UYGULANAB L RL HAZIRLAYAN Ö RENC LER: Tayanç HASANZADE Ahmet Rasim KARSLIO LU DANI MAN Ö RETMEN: Mesut ESEN
ÖZEL EGE L SES BAZI ISKARTA BALIKLARIN (Isparoz, Hani) ETLER NDEN ALTERNAT F GÜBRE YAPIMI VE UYGULANAB L RL HAZIRLAYAN Ö RENC LER: Tayanç HASANZADE Ahmet Rasim KARSLIO LU DANI MAN Ö RETMEN: Mesut ESEN
ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ
 ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ BATI AKDENİZ ÜZÜM ÇEŞİTLERİNİN MOLEKÜLER KARAKTERİZASYONU VE ÜLKE ASMA KAYNAKLARI İLE GENETİK İLİŞKİSİ Şirin ASLANTAŞ Danışman Öğretim Üyesi
ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ BATI AKDENİZ ÜZÜM ÇEŞİTLERİNİN MOLEKÜLER KARAKTERİZASYONU VE ÜLKE ASMA KAYNAKLARI İLE GENETİK İLİŞKİSİ Şirin ASLANTAŞ Danışman Öğretim Üyesi
ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ
 ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ MANİSA, İZMİR, AYDIN, MUĞLA VE KÜTAHYA İLLERİNE AİT ASMA GEN KAYNAKLARININ SSRs (Simple Sequence Repeats) A DAYALI GENETİK KARAKTERİZASYONU
ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ MANİSA, İZMİR, AYDIN, MUĞLA VE KÜTAHYA İLLERİNE AİT ASMA GEN KAYNAKLARININ SSRs (Simple Sequence Repeats) A DAYALI GENETİK KARAKTERİZASYONU
Bunlardan SSR (Simple Sequence Repeat=Basit dizi tekrarları)(mikrosatelit) markörler yüksek tanımlama kabiliyeti, tekrarlanma oranının çok yüksek oluş
(mikrosatelit) markörler yüksek tanımlama kabiliyeti, tekrarlanma oranının çok yüksek oluş") 1. GİRİŞ Asma türleri içerisinde en önemlisi olan Vitis vinifera L. türü, dünya üzüm üretiminin tamamına yakınını karşılamaktadır. Türün ilk olarak Kafkasya ve Anadolu da kültüre alındığı ve Anadolu nun
1. GİRİŞ Asma türleri içerisinde en önemlisi olan Vitis vinifera L. türü, dünya üzüm üretiminin tamamına yakınını karşılamaktadır. Türün ilk olarak Kafkasya ve Anadolu da kültüre alındığı ve Anadolu nun
SNP TEK NÜKLEOTİD POLİMORFİZMLERİ (SINGLE NUCLEOTIDE POLYMORPHISMS)
") SNP TEK NÜKLEOTİD POLİMORFİZMLERİ (SINGLE NUCLEOTIDE POLYMORPHISMS) Herhangi iki bireyin DNA dizisi %99.9 aynıdır. %0.1 = ~3x10 6 nükleotid farklılığı sağlar. Genetik materyalde varyasyon : Polimorfizm
SNP TEK NÜKLEOTİD POLİMORFİZMLERİ (SINGLE NUCLEOTIDE POLYMORPHISMS) Herhangi iki bireyin DNA dizisi %99.9 aynıdır. %0.1 = ~3x10 6 nükleotid farklılığı sağlar. Genetik materyalde varyasyon : Polimorfizm
Domuz Ayrığı (Dactylis glomerata L.) Populasyonlarında Genetik Çeşitliliğin Belirlenmesi
 Populasyonlarında Genetik Çeşitliliğin Belirlenmesi") Ordu Üniversitesi Bilim ve Teknoloji Dergisi / Ordu University Journal of Science and Technology Ordu Üniv. Bil. Tek. Derg., 2017; 7(2): 289-294 Ordu Univ. J. Sci. Tech., 2017; 7(2): 289-294 e-issn: 2146-6459
Ordu Üniversitesi Bilim ve Teknoloji Dergisi / Ordu University Journal of Science and Technology Ordu Üniv. Bil. Tek. Derg., 2017; 7(2): 289-294 Ordu Univ. J. Sci. Tech., 2017; 7(2): 289-294 e-issn: 2146-6459
POLİMERAZ ZİNCİR REAKSİYONU (PZR-PCR) VE RESTRİKSİYON PARÇA UZUNLUĞU POLİMORFİZMİ (RFLP)
 VE RESTRİKSİYON PARÇA UZUNLUĞU POLİMORFİZMİ (RFLP)") Deney: M 1 POLİMERAZ ZİNCİR REAKSİYONU (PZR-PCR) VE RESTRİKSİYON PARÇA UZUNLUĞU POLİMORFİZMİ (RFLP) a) PCR yöntemi uygulaması b) RPLF sonuçları değerlendirilmesi I. Araç ve Gereç dntp (deoksi Nükleotid
Deney: M 1 POLİMERAZ ZİNCİR REAKSİYONU (PZR-PCR) VE RESTRİKSİYON PARÇA UZUNLUĞU POLİMORFİZMİ (RFLP) a) PCR yöntemi uygulaması b) RPLF sonuçları değerlendirilmesi I. Araç ve Gereç dntp (deoksi Nükleotid
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
 ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ Ali SABIR BAZI ÜZÜM ÇEŞİT VE ANAÇLARININ AMPELOGRAFİK VE MOLEKÜLER KARAKTERİZASYONU BAHÇE BİTKİLERİ ANABİLİM DALI ADANA, 2008 ÇUKUROVA ÜNİVERSİTESİ
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ Ali SABIR BAZI ÜZÜM ÇEŞİT VE ANAÇLARININ AMPELOGRAFİK VE MOLEKÜLER KARAKTERİZASYONU BAHÇE BİTKİLERİ ANABİLİM DALI ADANA, 2008 ÇUKUROVA ÜNİVERSİTESİ
MELEZLEME ISLAHI İLE ELDE EDİLEN BAZI ÜZÜM ÇEŞİTLERİNİN EBEVEYN ANALİZLERİ VE ÇEKİRDEKSİZ FERTLERİN MARKÖRE DAYALI SELEKSİYONU
 MELEZLEME ISLAHI İLE ELDE EDİLEN BAZI ÜZÜM ÇEŞİTLERİNİN EBEVEYN ANALİZLERİ VE ÇEKİRDEKSİZ FERTLERİN MARKÖRE DAYALI SELEKSİYONU Ayça KARAUZ Doktora Tezi Bahçe Bitkileri Anabilim Dalı Danışman: Prof. Dr.
MELEZLEME ISLAHI İLE ELDE EDİLEN BAZI ÜZÜM ÇEŞİTLERİNİN EBEVEYN ANALİZLERİ VE ÇEKİRDEKSİZ FERTLERİN MARKÖRE DAYALI SELEKSİYONU Ayça KARAUZ Doktora Tezi Bahçe Bitkileri Anabilim Dalı Danışman: Prof. Dr.
I. Projenin Türkçe ve İngilizce Adı ve Özetleri İvesi Koyunlarında mikrosatellite lokuslarında polimorfizmin tespiti Güneydoğu Anadolu Tarımsal Araştı
 T.C. ANKARA ÜNİVERSİTESİ BİLİMSEL ARAŞTIRMA PROJESİ KESİN RAPORU İvesi Koyunlarında Mikrosatellite Lokuslarında Polimorfizmin Tespiti Proje Yürütücüsü: Profesör Doktor Ayhan ELİÇİN Proje Numarası: 20050711087
T.C. ANKARA ÜNİVERSİTESİ BİLİMSEL ARAŞTIRMA PROJESİ KESİN RAPORU İvesi Koyunlarında Mikrosatellite Lokuslarında Polimorfizmin Tespiti Proje Yürütücüsü: Profesör Doktor Ayhan ELİÇİN Proje Numarası: 20050711087
MOLEKÜLER BİYOLOJİ LABORATUVARI
 MOLEKÜLER 2014-2015 BİYOLOJİ LABORATUVARI GÜZ DÖNEMİ MOLEKÜLER BİYOLOJİ LABORATUVARI 7.HAFTA DERS NOTLARI GAZİ ÜNİVERSİTESİ FEN FAKÜLTESİ BİYOLOJİ BÖLÜMÜ Sayfa 1 / 6 1. RFLP (RESTRİKSİYON PARÇA UZUNLUK
MOLEKÜLER 2014-2015 BİYOLOJİ LABORATUVARI GÜZ DÖNEMİ MOLEKÜLER BİYOLOJİ LABORATUVARI 7.HAFTA DERS NOTLARI GAZİ ÜNİVERSİTESİ FEN FAKÜLTESİ BİYOLOJİ BÖLÜMÜ Sayfa 1 / 6 1. RFLP (RESTRİKSİYON PARÇA UZUNLUK
ÖZEL EGE LKÖ RET M OKULU DO RULARIN DANSI HAZIRLAYAN: YANKI TURGUT DANI MAN Ö RETMEN: AY EGÜL GÜRKAN
 ÖZEL EGE LKÖ RET M OKULU DO RULARIN DANSI HAZIRLAYAN: YANKI TURGUT DANI MAN Ö RETMEN: AY EGÜL GÜRKAN zmir,2009 1 TE EKKÜR Bu projede eme i geçen danı man ö retmenim Ay egül Gürkan a, sunum hazırlamamda
ÖZEL EGE LKÖ RET M OKULU DO RULARIN DANSI HAZIRLAYAN: YANKI TURGUT DANI MAN Ö RETMEN: AY EGÜL GÜRKAN zmir,2009 1 TE EKKÜR Bu projede eme i geçen danı man ö retmenim Ay egül Gürkan a, sunum hazırlamamda
T.H. Morgan ve A.H. Sturtevant 1911
 GENETĐK 111-503 HAFTA III Bağlantı, Asosiyasyon, Haritalama Doç. Dr. Hilâl Özdağ T.H. Morgan ve A.H. Sturtevant 1911 Morgan ın soruları: 1. Gen ayrılmasının kaynağı nedir? Janssens ve ark: kiyazma 2. Görünüşteki
GENETĐK 111-503 HAFTA III Bağlantı, Asosiyasyon, Haritalama Doç. Dr. Hilâl Özdağ T.H. Morgan ve A.H. Sturtevant 1911 Morgan ın soruları: 1. Gen ayrılmasının kaynağı nedir? Janssens ve ark: kiyazma 2. Görünüşteki
Sebze Islahında Moleküler Markırların Kullanımı
 Sebze Islahında Moleküler Markırların Kullanımı Esra CEBECİ Ziraat Yüksek Mühendisi 28.12.2012-28.06.2013 Atatürk Bahçe Kültürleri Merkez Araştırma Enstitüsü YALOVA Sunu Planı Çalışmanın tanıtımı, Yapılan
Sebze Islahında Moleküler Markırların Kullanımı Esra CEBECİ Ziraat Yüksek Mühendisi 28.12.2012-28.06.2013 Atatürk Bahçe Kültürleri Merkez Araştırma Enstitüsü YALOVA Sunu Planı Çalışmanın tanıtımı, Yapılan
BİLECİK ŞEYH EDEBALİ ÜNİVERSİTESİ AKADEMİK ÖZGEÇMİŞ FORMU
 Adı Soyadı Ünvanı Birimi Doğum Yeri/Yılı E-Posta BİLECİK ŞEYH EDEBALİ ÜNİVERSİTESİ AKADEMİK ÖZGEÇMİŞ FORMU KİŞİSEL BİLGİLER Özlem ÇALKAN SAĞLAM Yrd. Doç. Dr. Tarım Bilimleri ve Teknolojikleri Fakültesi
Adı Soyadı Ünvanı Birimi Doğum Yeri/Yılı E-Posta BİLECİK ŞEYH EDEBALİ ÜNİVERSİTESİ AKADEMİK ÖZGEÇMİŞ FORMU KİŞİSEL BİLGİLER Özlem ÇALKAN SAĞLAM Yrd. Doç. Dr. Tarım Bilimleri ve Teknolojikleri Fakültesi
ORMAN AĞACI ISLAHI. Yrd. Doç. Dr. DENİZ GÜNEY ( GÜZ DÖNEMİ)
") ORMAN AĞACI ISLAHI Yrd. Doç. Dr. DENİZ GÜNEY (2015-2016 GÜZ DÖNEMİ) Hızlı nüfus artışı, sanayi ve teknolojideki gelişmeler, küresel ısınmanın etkileriyle birleşerek ekosistem dengesi üzerinde yoğun baskı
ORMAN AĞACI ISLAHI Yrd. Doç. Dr. DENİZ GÜNEY (2015-2016 GÜZ DÖNEMİ) Hızlı nüfus artışı, sanayi ve teknolojideki gelişmeler, küresel ısınmanın etkileriyle birleşerek ekosistem dengesi üzerinde yoğun baskı
Perennial Identification of Erciş Grape Variety Clones
 YYÜ TAR BİL DERG (YYU J AGR SCI) 2011, 21(1):49-56 Geliş Tarihi (Received): 03.11.2010 Kabul Tarihi (Accepted): 24.12.2010 Araştırma Makalesi/Research Article (Original Paper) Erciş Üzüm Çeşidi Klonlarının
YYÜ TAR BİL DERG (YYU J AGR SCI) 2011, 21(1):49-56 Geliş Tarihi (Received): 03.11.2010 Kabul Tarihi (Accepted): 24.12.2010 Araştırma Makalesi/Research Article (Original Paper) Erciş Üzüm Çeşidi Klonlarının
HAFTA III Bağlantı, Asosiyasyon, Haritalama
 Biyoteknoloji ve Genetik I HAFTA III Bağlantı, Asosiyasyon, Haritalama Prof. Dr. Hilâl Özdağ T.H. Morgan ve A.H. Sturtevant 1911 Morgan ın soruları: 1. Gen ayrılmasının kaynağı nedir? Janssens ve ark:
Biyoteknoloji ve Genetik I HAFTA III Bağlantı, Asosiyasyon, Haritalama Prof. Dr. Hilâl Özdağ T.H. Morgan ve A.H. Sturtevant 1911 Morgan ın soruları: 1. Gen ayrılmasının kaynağı nedir? Janssens ve ark:
BAZI KARPUZ GENOTİPLERİNİN SSR ve SRAP MARKÖRLERİ ile KARAKTERİZASYONU 1. Characterization of Some Watermelon Genotypes by SSR and SRAP Markers
 BAZI KARPUZ GENOTİPLERİNİN SSR ve SRAP MARKÖRLERİ ile KARAKTERİZASYONU 1 Characterization of Some Watermelon Genotypes by SSR and SRAP Markers İlknur SOLMAZ Bahçe Bitkileri Anabilim Dalı Nebahat SARI Bahçe
BAZI KARPUZ GENOTİPLERİNİN SSR ve SRAP MARKÖRLERİ ile KARAKTERİZASYONU 1 Characterization of Some Watermelon Genotypes by SSR and SRAP Markers İlknur SOLMAZ Bahçe Bitkileri Anabilim Dalı Nebahat SARI Bahçe
atak@yalovabahce.gov.tr /atakarif@gmail.com Ege Üniversitesi Fen Bilimleri Enstitüsü Bahçe Bitkileri Anabilim Dalı / 2010
 KİŞİSEL BİLGİLER Adı Soyadı Dr. Arif ATAK Unvan Dr.Mühendis Telefon 0226 814 25 20 (1220) E-mail atak@yalovabahce.gov.tr /atakarif@gmail.com Doğum Tarihi - Yeri 15.11.1972 Aksaray Fotoğraf Doktora Üniversite
KİŞİSEL BİLGİLER Adı Soyadı Dr. Arif ATAK Unvan Dr.Mühendis Telefon 0226 814 25 20 (1220) E-mail atak@yalovabahce.gov.tr /atakarif@gmail.com Doğum Tarihi - Yeri 15.11.1972 Aksaray Fotoğraf Doktora Üniversite
YÜKSEKÖĞRETİM KURULU YARDIMCI DOÇENT 01.12.2014. : Sinop Üniversitesi Fen Edebiyat Fakültesi Biyoloji Bölümü Sinop
 HÜLYA SİPAHİ ÖZGEÇMİŞ YÜKSEKÖĞRETİM KURULU YARDIMCI DOÇENT 01.12.2014 Adres : Sinop Üniversitesi Fen Edebiyat Fakültesi Biyoloji Bölümü Sinop Telefon : 3682715516-4206 E-posta Doğum Tarihi : Faks : Kadro
HÜLYA SİPAHİ ÖZGEÇMİŞ YÜKSEKÖĞRETİM KURULU YARDIMCI DOÇENT 01.12.2014 Adres : Sinop Üniversitesi Fen Edebiyat Fakültesi Biyoloji Bölümü Sinop Telefon : 3682715516-4206 E-posta Doğum Tarihi : Faks : Kadro
ASMAGİLLER Asmalar; Rhamnales
 ASMAGİLLER Asmalar; Rhamnales takımında yer alan, sarılıcı gövde yapısına sahip bitkilerin yanısıra, küçük ağaç ve çalı formundaki türlerden oluşan bitkilerdir. A S M A : Vitis cinsinin sistematiği Takım
ASMAGİLLER Asmalar; Rhamnales takımında yer alan, sarılıcı gövde yapısına sahip bitkilerin yanısıra, küçük ağaç ve çalı formundaki türlerden oluşan bitkilerdir. A S M A : Vitis cinsinin sistematiği Takım
TÜRKİYE DE BULUNAN BAZI YERLİ SIĞIR IRKLARININ GENETİK YAPILARININ KARAKTERİZASYONU
 TÜRKİYE DE BULUNAN BAZI YERLİ SIĞIR IRKLARININ GENETİK YAPILARININ KARAKTERİZASYONU DOKTORA TEZİ ZOOTEKNİ ANABİLİM DALI Danışman Yard. Doç. Dr. Ercan KURAR KONYA - 2011 TÜRKİYE DE BULUNAN BAZI YERLİ SIĞIR
TÜRKİYE DE BULUNAN BAZI YERLİ SIĞIR IRKLARININ GENETİK YAPILARININ KARAKTERİZASYONU DOKTORA TEZİ ZOOTEKNİ ANABİLİM DALI Danışman Yard. Doç. Dr. Ercan KURAR KONYA - 2011 TÜRKİYE DE BULUNAN BAZI YERLİ SIĞIR
Populasyon Genetiği. Populasyonlardaki alel ve gen frekanslarının değişmesine neden olan süreçleri araştıran evrimsel bilim dalı.
 Bu dersin içeriği, Populasyonun tanımı, Alel ve genotip frekansı, Gen havuzu, Gen frekansı, Gerçek/Doğal populasyonlar ve ideal populasyonlar, Populasyon genetiğinin çalışma alanları, HW kanunu -giriş,
Bu dersin içeriği, Populasyonun tanımı, Alel ve genotip frekansı, Gen havuzu, Gen frekansı, Gerçek/Doğal populasyonlar ve ideal populasyonlar, Populasyon genetiğinin çalışma alanları, HW kanunu -giriş,
Genom analizi için belirteç olarak kullanılan DNA dizileri
 Salgınların Araştırılmasında Hızlı Genotiplendirme Yöntemleri Avantajları-Dezavantajları Doç. Dr. Z. Ceren KARAHAN Ankara Üniversitesi Tıp Fakültesi Tıbbi Mikrobioyoloji Anabilim Dalı Moleküler Genetik
Salgınların Araştırılmasında Hızlı Genotiplendirme Yöntemleri Avantajları-Dezavantajları Doç. Dr. Z. Ceren KARAHAN Ankara Üniversitesi Tıp Fakültesi Tıbbi Mikrobioyoloji Anabilim Dalı Moleküler Genetik
YÖNETMELİK KAFKAS ÜNİVERSİTESİ ARICILIĞI GELİŞTİRME UYGULAMA VE ARAŞTIRMA MERKEZİ YÖNETMELİĞİ
 22 Mayıs 2012 SALI Resmî Gazete Sayı : 28300 Kafkas Üniversitesinden: YÖNETMELİK KAFKAS ÜNİVERSİTESİ ARICILIĞI GELİŞTİRME UYGULAMA VE ARAŞTIRMA MERKEZİ YÖNETMELİĞİ BİRİNCİ BÖLÜM Amaç, Kapsam, Dayanak ve
22 Mayıs 2012 SALI Resmî Gazete Sayı : 28300 Kafkas Üniversitesinden: YÖNETMELİK KAFKAS ÜNİVERSİTESİ ARICILIĞI GELİŞTİRME UYGULAMA VE ARAŞTIRMA MERKEZİ YÖNETMELİĞİ BİRİNCİ BÖLÜM Amaç, Kapsam, Dayanak ve
PERMEAB L TE VE KONSOL DASYON DENEYLER LE ELDE ED LEN PERMEAB L TE KATSAYILARININ KAR ILA TIRILMASI
 PERMEAB L TE VE KONSOL DASYON DENEYLER LE ELDE ED LEN PERMEAB L TE KATSAYILARININ KAR ILA TIRILMASI Sami ARSOY Erdinç KESK N Cüneyt YILMAZ Doç. Dr. Ar.Gör. Ar.Gör. Kocaeli Üniversitesi Kocaeli Üniversitesi
PERMEAB L TE VE KONSOL DASYON DENEYLER LE ELDE ED LEN PERMEAB L TE KATSAYILARININ KAR ILA TIRILMASI Sami ARSOY Erdinç KESK N Cüneyt YILMAZ Doç. Dr. Ar.Gör. Ar.Gör. Kocaeli Üniversitesi Kocaeli Üniversitesi
SALGIN ARAŞTIRMASINDA KULLANILAN TİPLENDİRME YÖNTEMLERİ
 SALGIN ARAŞTIRMASINDA KULLANILAN TİPLENDİRME YÖNTEMLERİ Prof.Dr. Meltem Yalınay Çırak Gazi Üniversitesi Tıp Fakültesi Mikrobiyoloji ve Klinik Mikrobiyoloji A.D. fenotipik yöntemler genotipik yöntemler
SALGIN ARAŞTIRMASINDA KULLANILAN TİPLENDİRME YÖNTEMLERİ Prof.Dr. Meltem Yalınay Çırak Gazi Üniversitesi Tıp Fakültesi Mikrobiyoloji ve Klinik Mikrobiyoloji A.D. fenotipik yöntemler genotipik yöntemler
Key words: Transport sector, greenhouse gas emissions, road transport
 mından Kaynaklanan Sera Gazı Emisyonları n erlendirilmesi l Diler 1, bay 3 n Özet Bu çalı mada iklim de i ikli ine neden olan, karayolu ula ımından kaynaklanan sera gazı emisyonları ülke bazında, bölgesel
mından Kaynaklanan Sera Gazı Emisyonları n erlendirilmesi l Diler 1, bay 3 n Özet Bu çalı mada iklim de i ikli ine neden olan, karayolu ula ımından kaynaklanan sera gazı emisyonları ülke bazında, bölgesel
DR. ONUR YILMAZ. Adnan Menderes Üniversitesi Ziraat Fakültesi Zootekni Bölümü Biyometri & Genetik A.B.D.
 DR. ONUR YILMAZ Adnan Menderes Üniversitesi Ziraat Fakültesi Zootekni Bölümü Biyometri & Genetik A.B.D. Koç Katımı Çiftleşme mevsiminde kızgınlık gösteren koyunun koç ile birleştirilmesi olayına koç katımı
DR. ONUR YILMAZ Adnan Menderes Üniversitesi Ziraat Fakültesi Zootekni Bölümü Biyometri & Genetik A.B.D. Koç Katımı Çiftleşme mevsiminde kızgınlık gösteren koyunun koç ile birleştirilmesi olayına koç katımı
BİLGİSAYAR PROGRAMLARI YARDIMIYLA ŞEV DURAYLILIK ANALİZLERİ * Software Aided Slope Stability Analysis*
 BİLGİSAYAR PROGRAMLARI YARDIMIYLA ŞEV DURAYLILIK ANALİZLERİ * Software Aided Slope Stability Analysis* Mustafa Özgür KESKİN Maden Mühendisliği Anabilim Dalı Ahmet M. KILIÇ Maden Mühendisliği Anabilim Dalı
BİLGİSAYAR PROGRAMLARI YARDIMIYLA ŞEV DURAYLILIK ANALİZLERİ * Software Aided Slope Stability Analysis* Mustafa Özgür KESKİN Maden Mühendisliği Anabilim Dalı Ahmet M. KILIÇ Maden Mühendisliği Anabilim Dalı
ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ
 ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ DOĞU ANADOLU DA YETİŞEN BAZI ARMUT GEN KAYNAKLARININ SSR s(simple Sequence Repeats) MARKÖRLERE DAYALI GENETİK KARAKTERİZASYONU Hatice ALTINBAY
ANKARA ÜNİVERSİTESİ BİYOTEKNOLOJİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ DOĞU ANADOLU DA YETİŞEN BAZI ARMUT GEN KAYNAKLARININ SSR s(simple Sequence Repeats) MARKÖRLERE DAYALI GENETİK KARAKTERİZASYONU Hatice ALTINBAY
I. Sabit Kaynaklar, bunlar ısınma ve üretim amaçlı faaliyetlerin yapıldı ı yerlerdir.
 G R Havanın gerek insan sa lı ına, gerekse do aya zarar verici duruma gelmesi, kirletici denen unsurların fazlala masıyla olur. Kirleticiler, belirli bir kaynaktan atmosfere bırakılan birinci derecede
G R Havanın gerek insan sa lı ına, gerekse do aya zarar verici duruma gelmesi, kirletici denen unsurların fazlala masıyla olur. Kirleticiler, belirli bir kaynaktan atmosfere bırakılan birinci derecede
Derece Bölüm/Program Üniversite l. Doktora Bahçe Bitkileri Ankara Üniversitesi 2008
 ÖZGEÇM VE YAYIN L STES ÖZGEÇM Ad Soyad : Volkan GÖZEN Do um Tarihi ve Yeri: 20 Nisan 1973, Malatya Medeni Durumu: Evli REN M DURUMU Derece Bölüm/Program Üniversite l Lisans Tar msal Yap lar ve Ege Üniversitesi
ÖZGEÇM VE YAYIN L STES ÖZGEÇM Ad Soyad : Volkan GÖZEN Do um Tarihi ve Yeri: 20 Nisan 1973, Malatya Medeni Durumu: Evli REN M DURUMU Derece Bölüm/Program Üniversite l Lisans Tar msal Yap lar ve Ege Üniversitesi
Söke İlçesinde Pnömatik Ekim Makinaları Talep Projeksiyonunun Belirlenmesi*
 91 Söke İlçesinde Pnömatik Ekim Makinaları Talep Projeksiyonunun Belirlenmesi* Hakan Destici (1) Cengiz Özarslan (2) (1) Söke Ziraat Odası, Söke / Aydın (2) ADÜ Ziraat Fakültesi Tarım Makinaları Bölümü,
91 Söke İlçesinde Pnömatik Ekim Makinaları Talep Projeksiyonunun Belirlenmesi* Hakan Destici (1) Cengiz Özarslan (2) (1) Söke Ziraat Odası, Söke / Aydın (2) ADÜ Ziraat Fakültesi Tarım Makinaları Bölümü,
Parkinson Hastalığı ile α-sinüklein Geni Polimorfizmlerinin İlişkisinin Araştırılması
 İ.Ü. CERRAHPAŞA TIP FAKÜLTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ ANABİLİM DALI Parkinson Hastalığı ile α-sinüklein Geni Polimorfizmlerinin İlişkisinin Araştırılması Araş.Gör. Yener KURMAN İSTANBUL
İ.Ü. CERRAHPAŞA TIP FAKÜLTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ ANABİLİM DALI Parkinson Hastalığı ile α-sinüklein Geni Polimorfizmlerinin İlişkisinin Araştırılması Araş.Gör. Yener KURMAN İSTANBUL
TÜBİTAK BİDEB LİSE ÖĞRETMENLERİ FİZİK, KİMYA, BİYOLOJİ, MATEMATİK- PROJE DANIŞMANLIĞI EĞİTİMİ ÇALIŞTAYI LİSE3 (Çalıştay 2013) BİYOLOJİ GRUP TUHAF
 BİYOLOJİ GRUP TUHAF") TÜBİTAK BİDEB LİSE ÖĞRETMENLERİ FİZİK, KİMYA, BİYOLOJİ, MATEMATİK- PROJE DANIŞMANLIĞI EĞİTİMİ ÇALIŞTAYI LİSE3 (Çalıştay 2013) BİYOLOJİ GRUP TUHAF PROJE ÖNERİSİ ADI TUHAF MATERYALLERDEN İZOLE EDİLEN DNA
TÜBİTAK BİDEB LİSE ÖĞRETMENLERİ FİZİK, KİMYA, BİYOLOJİ, MATEMATİK- PROJE DANIŞMANLIĞI EĞİTİMİ ÇALIŞTAYI LİSE3 (Çalıştay 2013) BİYOLOJİ GRUP TUHAF PROJE ÖNERİSİ ADI TUHAF MATERYALLERDEN İZOLE EDİLEN DNA
Gen Arama Yordamı ve Nörolojik Hastalıklarla İlgili Gen Keşfi Çalışmalarına Türkiye den Örnekler
 Gen Arama Yordamı ve Nörolojik Hastalıklarla İlgili Gen Keşfi Çalışmalarına Türkiye den Örnekler Doç. Dr. Sibel Aylin Uğur İstanbul Üniversitesi Deneysel Tıp Araştırma Enstitüsü-Genetik 13. Ulusal Sinirbilim
Gen Arama Yordamı ve Nörolojik Hastalıklarla İlgili Gen Keşfi Çalışmalarına Türkiye den Örnekler Doç. Dr. Sibel Aylin Uğur İstanbul Üniversitesi Deneysel Tıp Araştırma Enstitüsü-Genetik 13. Ulusal Sinirbilim
1. EKSİK BASKINLIK 2. EŞ BASKINLIK 3. ÇOK ALLELLİLİK 4. KONTROL ÇAPRAZLAMASI
 1. EKSİK BASKINLIK 2. EŞ BASKINLIK 3. ÇOK ALLELLİLİK 4. KONTROL ÇAPRAZLAMASI Eksik baskınlık (ekivalentlik): Bazı karakterlerde allel genler, birbirine tam baskınlık göstermezler. Bireyler, bu karakterler
1. EKSİK BASKINLIK 2. EŞ BASKINLIK 3. ÇOK ALLELLİLİK 4. KONTROL ÇAPRAZLAMASI Eksik baskınlık (ekivalentlik): Bazı karakterlerde allel genler, birbirine tam baskınlık göstermezler. Bireyler, bu karakterler
DNA Dizileme (Sekanslama)
") T.C GIDA TARIM VE HAYVANCILIK BAKANLIĞI PENDİK VETERİNER KONTROL ENSTİTÜSÜ DNA Dizileme (Sekanslama) Dr. Eray ATIL Vet. Hekim, Mikrobiyolog Pendik Veteriner Kontrol Enstitüsü Eğitim Bilgileri Eğitim süresi
T.C GIDA TARIM VE HAYVANCILIK BAKANLIĞI PENDİK VETERİNER KONTROL ENSTİTÜSÜ DNA Dizileme (Sekanslama) Dr. Eray ATIL Vet. Hekim, Mikrobiyolog Pendik Veteriner Kontrol Enstitüsü Eğitim Bilgileri Eğitim süresi
BİLECİK ŞEYH EDEBALİ ÜNİVERSİTESİ AKADEMİK ÖZGEÇMİŞ FORMU
 Adı Soyadı Ünvanı Birimi Doğum Yeri/Yılı E-Posta BİLECİK ŞEYH EDEBALİ ÜNİVERSİTESİ AKADEMİK ÖZGEÇMİŞ FORMU KİŞİSEL BİLGİLER Hayri SAĞLAM Yrd. Doç. Dr. Ziraat ve Doğa Bilimleri Fakültesi Bahçe Bitkileri
Adı Soyadı Ünvanı Birimi Doğum Yeri/Yılı E-Posta BİLECİK ŞEYH EDEBALİ ÜNİVERSİTESİ AKADEMİK ÖZGEÇMİŞ FORMU KİŞİSEL BİLGİLER Hayri SAĞLAM Yrd. Doç. Dr. Ziraat ve Doğa Bilimleri Fakültesi Bahçe Bitkileri
TARIMSAL BİYOTEKNOLOJİYE GİRİŞ
 TARIMSAL BİYOTEKNOLOJİYE GİRİŞ Bitki Doku Kültürü Adnan Menderes Üniversitesi Tarımsal Biyoteknoloji Bölümü TB101 Çiğdem Yamaner (Yrd. Doç. Dr.) 4. Hafta (08.10.2013) ADÜ Tarımsal Biyoteknoloji Bölümü
TARIMSAL BİYOTEKNOLOJİYE GİRİŞ Bitki Doku Kültürü Adnan Menderes Üniversitesi Tarımsal Biyoteknoloji Bölümü TB101 Çiğdem Yamaner (Yrd. Doç. Dr.) 4. Hafta (08.10.2013) ADÜ Tarımsal Biyoteknoloji Bölümü
AİLESEL AKDENİZ ATEŞİ (AAA-FMF)
") AİLESEL AKDENİZ ATEŞİ (AAA-FMF) MOLEKÜLER YAKLAŞIMLAR DÜZEN GENETİK HASTALIKLAR TANI MERKEZİ SERPİL ERASLAN, PhD AİLESEL AKDENİZ ATEŞİ Otozomal resesif kalıtım Akdeniz ve Ortadoğu kökenli populasyonlarda
AİLESEL AKDENİZ ATEŞİ (AAA-FMF) MOLEKÜLER YAKLAŞIMLAR DÜZEN GENETİK HASTALIKLAR TANI MERKEZİ SERPİL ERASLAN, PhD AİLESEL AKDENİZ ATEŞİ Otozomal resesif kalıtım Akdeniz ve Ortadoğu kökenli populasyonlarda
MOLEKÜLER TANISI DÜZEN GENETİK HASTALIKLAR TANI MERKEZİ. SERPİL ERASLAN, PhD
 β-talaseminin MOLEKÜLER TANISI DÜZEN GENETİK HASTALIKLAR TANI MERKEZİ SERPİL ERASLAN, PhD BETA TALASEMİ HEMOGLOBİNOPATİLER Otozomal resesif (globin gen ailesi) Özellikle Çukurova, Akdeniz kıyı şeridi,
β-talaseminin MOLEKÜLER TANISI DÜZEN GENETİK HASTALIKLAR TANI MERKEZİ SERPİL ERASLAN, PhD BETA TALASEMİ HEMOGLOBİNOPATİLER Otozomal resesif (globin gen ailesi) Özellikle Çukurova, Akdeniz kıyı şeridi,
Mutasyon: DNA dizisinde meydana gelen kalıcı değişiklik. Polimorfizm: iki veya daha fazla farklı fenotipin aynı tür popülasyonunda bulunmasıdır.
 Allel: Bir genin seçenekli biçimi Wild Tip: Normal allel. Bireylerin çoğunda bulunan Mutasyon: DNA dizisinde meydana gelen kalıcı değişiklik Polimorfizm: iki veya daha fazla farklı fenotipin aynı tür popülasyonunda
Allel: Bir genin seçenekli biçimi Wild Tip: Normal allel. Bireylerin çoğunda bulunan Mutasyon: DNA dizisinde meydana gelen kalıcı değişiklik Polimorfizm: iki veya daha fazla farklı fenotipin aynı tür popülasyonunda
hendisliği BYM613 Genetik MühendisliM Tanımlar: Gen, genom DNA ve yapısı, Nükleik asitler Genetik şifre DNA replikasyonu
 BYM613 Genetik MühendisliM hendisliği Hacettepe Üniversitesi Biyomühendislik BölümüB 2012-2013 2013 Güz G z DönemiD Salı 9.00-11.45, D9 Dr. Eda Çelik-AKDUR edacelik@hacettepe.edu.tr İçerik Tanımlar: Gen,
BYM613 Genetik MühendisliM hendisliği Hacettepe Üniversitesi Biyomühendislik BölümüB 2012-2013 2013 Güz G z DönemiD Salı 9.00-11.45, D9 Dr. Eda Çelik-AKDUR edacelik@hacettepe.edu.tr İçerik Tanımlar: Gen,
Bornova Misketi Ve Cabernet Sauvignon Üzüm Çeşitlerinde Organik Ve Konvansiyonel Yetiştiriciliğin Asmanın Gelişimine, Üzüm Ve Şarap Kalitesine Etkisi
 Türkiye IV. Organik Tarım Sempozyumu, 28 Haziran - 1 Temmuz 2010, Erzurum, (Sunulu Bildiri) Bornova Misketi Ve Cabernet Sauvignon Üzüm Çeşitlerinde Organik Ve Konvansiyonel Yetiştiriciliğin Asmanın Gelişimine,
Türkiye IV. Organik Tarım Sempozyumu, 28 Haziran - 1 Temmuz 2010, Erzurum, (Sunulu Bildiri) Bornova Misketi Ve Cabernet Sauvignon Üzüm Çeşitlerinde Organik Ve Konvansiyonel Yetiştiriciliğin Asmanın Gelişimine,
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
 ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ Meryem Aylin AKYÜZ ANTEPFISTIĞINDA SİİRT X BAĞYOLU F1 POPULASYONU KULLANILARAK SSR MARKÖRLERI İLE GENETİK HARİTALAMA BİYOTEKNOLOJİ ANABİLİM
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ Meryem Aylin AKYÜZ ANTEPFISTIĞINDA SİİRT X BAĞYOLU F1 POPULASYONU KULLANILARAK SSR MARKÖRLERI İLE GENETİK HARİTALAMA BİYOTEKNOLOJİ ANABİLİM
ASMADA QTL (KANTİTATİF KARAKTER LOKUS) ANALİZİ. Burçak İŞÇİ. Ege Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümü Bornova-İzmir/TURKEY
 ANALİZİ. Burçak İŞÇİ. Ege Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümü Bornova-İzmir/TURKEY") ANADOLU, J. of AARI 18 (2) 2008, 11 37 MARA ASMADA QTL (KANTİTATİF KARAKTER LOKUS) ANALİZİ Burçak İŞÇİ Ege Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümü Bornova-İzmir/TURKEY ÖZ: Son yıllarda, DNA
ANADOLU, J. of AARI 18 (2) 2008, 11 37 MARA ASMADA QTL (KANTİTATİF KARAKTER LOKUS) ANALİZİ Burçak İŞÇİ Ege Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümü Bornova-İzmir/TURKEY ÖZ: Son yıllarda, DNA
ORMAN AĞACI ISLAHI. Yrd. Doç. Dr. DENİZ GÜNEY ( GÜZ DÖNEMİ)
") ORMAN AĞACI ISLAHI Yrd. Doç. Dr. DENİZ GÜNEY (2015-2016 GÜZ DÖNEMİ) MELEZ VE MELEZ GÜCÜ (HETEROSİS): Kalıtsal özellikleri farklı iki bireyin çaprazlanması olayına "Melezleme" denir. Melezleme sonunda ortaya
ORMAN AĞACI ISLAHI Yrd. Doç. Dr. DENİZ GÜNEY (2015-2016 GÜZ DÖNEMİ) MELEZ VE MELEZ GÜCÜ (HETEROSİS): Kalıtsal özellikleri farklı iki bireyin çaprazlanması olayına "Melezleme" denir. Melezleme sonunda ortaya
1. Sınıf Güz Dönemi I. Hafta Pazartesi Salı Çarşamba Perşembe Cuma Ders Saati
 I. Hafta Ders Saati 15.09.2014 16.09.2014 17.09.2014 18.09.2014 19.09.2014 Atatürk İlkeleri ve İnkılap Tarihi I: Atatürk İlkeleri ve İnkılap Tarihi I: Makromoleküller (Yrd. Doç. Dr. Mehmet Ataş) Türk Dili
I. Hafta Ders Saati 15.09.2014 16.09.2014 17.09.2014 18.09.2014 19.09.2014 Atatürk İlkeleri ve İnkılap Tarihi I: Atatürk İlkeleri ve İnkılap Tarihi I: Makromoleküller (Yrd. Doç. Dr. Mehmet Ataş) Türk Dili
GENETİK POLİMORFİZMLER. Prof. Dr. Filiz ÖZBAŞ GERÇEKER
 GENETİK POLİMORFİZMLER Prof. Dr. Filiz ÖZBAŞ GERÇEKER Genomu bir kitap olarak düşünürsek... (Ridley, 2000) Kromozom olarak adlandırılan 23 bölüm Her bölüm birkaç bin hikayeden oluşur ki bunlar genlerdir.
GENETİK POLİMORFİZMLER Prof. Dr. Filiz ÖZBAŞ GERÇEKER Genomu bir kitap olarak düşünürsek... (Ridley, 2000) Kromozom olarak adlandırılan 23 bölüm Her bölüm birkaç bin hikayeden oluşur ki bunlar genlerdir.
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
 ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ Yüşa TÜRKELİ Pistacia vera L. X Pistacia atlantica Desf. MELEZ POPULASYONUNDA GENETİK HARİTALAMA BAHÇE BİTKİLERİ ANABİLİM DALI ADANA, 2010 ÇUKUROVA
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ Yüşa TÜRKELİ Pistacia vera L. X Pistacia atlantica Desf. MELEZ POPULASYONUNDA GENETİK HARİTALAMA BAHÇE BİTKİLERİ ANABİLİM DALI ADANA, 2010 ÇUKUROVA
10.Sınıf Biyoloji. Genetik. cevap anahtarı
 10.Sınıf Biyoloji 4 Genetik cevap anahtarı 4 1 KALITIM Canlı bireylere ait olan özelliklerin, yavru bireylere aktarılmasını inceleyen bilim dalına kalıtım denir. Aristo m.ö. 350 yılında kalıtımın kan yoluyla
10.Sınıf Biyoloji 4 Genetik cevap anahtarı 4 1 KALITIM Canlı bireylere ait olan özelliklerin, yavru bireylere aktarılmasını inceleyen bilim dalına kalıtım denir. Aristo m.ö. 350 yılında kalıtımın kan yoluyla
DNA TİPLEME YÖNTEMLERİ (SOMATİK STRLOKUSLARI)
") Adli Biyolojik Incelemeler ve Molekuler Genetik Kursu DNA TİPLEME YÖNTEMLERİ (SOMATİK STRLOKUSLARI) Yrd. Doç. Dr. GÖNÜL FİLOĞLU İ.Ü. ADLİ TIP ENSTİTÜSÜ KRİMİNAL İDANTİFİKASYONDA DNA TİPLEMESİ Moleküler
Adli Biyolojik Incelemeler ve Molekuler Genetik Kursu DNA TİPLEME YÖNTEMLERİ (SOMATİK STRLOKUSLARI) Yrd. Doç. Dr. GÖNÜL FİLOĞLU İ.Ü. ADLİ TIP ENSTİTÜSÜ KRİMİNAL İDANTİFİKASYONDA DNA TİPLEMESİ Moleküler
Bazı Ceviz (Juglans regia L.) Çeşitlerinin Çimlenme ve Çöğür (Anaçlık) Gelişme Performanslarının Belirlenmesi
 Çeşitlerinin Çimlenme ve Çöğür (Anaçlık) Gelişme Performanslarının Belirlenmesi") Bazı Ceviz (Juglans regia L.) Çeşitlerinin Çimlenme ve Çöğür (Anaçlık) Gelişme Performanslarının Belirlenmesi Akide ÖZCAN 1 Mehmet SÜTYEMEZ 2 1 Kahramanmaraş Sütçü İmam Üniv., Afşin Meslek Yüksekokulu,
Bazı Ceviz (Juglans regia L.) Çeşitlerinin Çimlenme ve Çöğür (Anaçlık) Gelişme Performanslarının Belirlenmesi Akide ÖZCAN 1 Mehmet SÜTYEMEZ 2 1 Kahramanmaraş Sütçü İmam Üniv., Afşin Meslek Yüksekokulu,
TÜBİTAK 1003 Buğday Tuzluluğu Projesinin Üçüncü Dönem Raporu Özeti
 TÜBİTAK 1003 Buğday Tuzluluğu Projesinin Üçüncü Dönem Raporu Özeti Toprak tuzluluğu, özellikle kurak ve yarı kurak bölgelerde buğday verimliliğini etkileyen başlıca tarımsal sorunlardan biridir. Ayrıca,
TÜBİTAK 1003 Buğday Tuzluluğu Projesinin Üçüncü Dönem Raporu Özeti Toprak tuzluluğu, özellikle kurak ve yarı kurak bölgelerde buğday verimliliğini etkileyen başlıca tarımsal sorunlardan biridir. Ayrıca,
Levent KESKİN 1 Mustafa PAKSOY 2,3 (paksoy42@hotmail.com) Önder TÜRKMEN 2. Bişkek/KIRGIZİSTAN
 Önder TÜRKMEN 2. Bişkek/KIRGIZİSTAN") Levent KESKİN Mustafa PAKSOY 2,3 (paksoy42@hotmail.com) Önder TÜRKMEN 2 :Batı Akdeniz Tarımsal Araştırmalar Enstitüsü Antalya/TÜRKİYE 2 :Selçuk Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümü Konya/TÜRKİYE
Levent KESKİN Mustafa PAKSOY 2,3 (paksoy42@hotmail.com) Önder TÜRKMEN 2 :Batı Akdeniz Tarımsal Araştırmalar Enstitüsü Antalya/TÜRKİYE 2 :Selçuk Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümü Konya/TÜRKİYE
TEMEL VETERĠNER GENETĠK
 DİKKATİNİZE: BURADA SADECE ÖZETİN İLK ÜNİTESİ SİZE ÖRNEK OLARAK GÖSTERİLMİŞTİR. ÖZETİN TAMAMININ KAÇ SAYFA OLDUĞUNU ÜNİTELERİ İÇİNDEKİLER BÖLÜMÜNDEN GÖREBİLİRSİNİZ. TEMEL VETERĠNER GENETĠK KISA ÖZET KOLAYAOF
DİKKATİNİZE: BURADA SADECE ÖZETİN İLK ÜNİTESİ SİZE ÖRNEK OLARAK GÖSTERİLMİŞTİR. ÖZETİN TAMAMININ KAÇ SAYFA OLDUĞUNU ÜNİTELERİ İÇİNDEKİLER BÖLÜMÜNDEN GÖREBİLİRSİNİZ. TEMEL VETERĠNER GENETĠK KISA ÖZET KOLAYAOF
ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ
 ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ BAZI STANDART ve KİLİS İLİ ZEYTİN (Olea europaea L.) ÇEŞİTLERİNİN SSRs (Simple Sequence Repeats) MARKÖRLER ARACILIĞIYLA GENETİK TANIMLANMASI
ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ BAZI STANDART ve KİLİS İLİ ZEYTİN (Olea europaea L.) ÇEŞİTLERİNİN SSRs (Simple Sequence Repeats) MARKÖRLER ARACILIĞIYLA GENETİK TANIMLANMASI
REKOMBİNANT DNA TEKNOLOJİSİ. Araş. Gör. Dr. Öğünç MERAL
 Araş. Gör. Dr. Öğünç MERAL 1960 lardan bu yana genetik ve moleküler biyolojideki kavrayışımızın hızla artması, biyoteknolojide heyecan verici buluşlar ve uygulamalara yol açtı. DNA yapısı ve fonksiyonlarının
Araş. Gör. Dr. Öğünç MERAL 1960 lardan bu yana genetik ve moleküler biyolojideki kavrayışımızın hızla artması, biyoteknolojide heyecan verici buluşlar ve uygulamalara yol açtı. DNA yapısı ve fonksiyonlarının
 Çiftlik hayvanları endüstrisinin yapısı elit Çok yönlü ticari Kantitatif genetik formulleri özeti Temel genetik: Genel öneri: Genellikle iki yönlü tablo kullanılır Sorular sorudaki probleme ilişkin verilen
Çiftlik hayvanları endüstrisinin yapısı elit Çok yönlü ticari Kantitatif genetik formulleri özeti Temel genetik: Genel öneri: Genellikle iki yönlü tablo kullanılır Sorular sorudaki probleme ilişkin verilen
Ara tırma Yöntem ve Teknikleri. Ay e Cabi
 Ara tırma Yöntem ve Teknikleri Ay e Cabi Bilim Nedir? Evreni anlama ve açıklama gayretlerinin tümüdür Soru sormakla ba lar Gerçe i anlama etkinlikleridir Organize edilmi bilgiler toplulu udur Metodik ve
Ara tırma Yöntem ve Teknikleri Ay e Cabi Bilim Nedir? Evreni anlama ve açıklama gayretlerinin tümüdür Soru sormakla ba lar Gerçe i anlama etkinlikleridir Organize edilmi bilgiler toplulu udur Metodik ve
B YOTEKNOLOJ ANAB L M DALI
 ÇUKUROVA ÜN VERS TES FEN B L MLER ENST TÜSÜ YÜKSEK L SANS TEZ Serkan AKCAN KAMAN CEV ZLER NDE APOM KS S OLASILI ININ MOLEKÜLER YÖNTEMLERLE ARA TIRILMASI B YOTEKNOLOJ ANAB L M DALI ADANA, 2007 ÇUKUROVA
ÇUKUROVA ÜN VERS TES FEN B L MLER ENST TÜSÜ YÜKSEK L SANS TEZ Serkan AKCAN KAMAN CEV ZLER NDE APOM KS S OLASILI ININ MOLEKÜLER YÖNTEMLERLE ARA TIRILMASI B YOTEKNOLOJ ANAB L M DALI ADANA, 2007 ÇUKUROVA
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
 ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ Burcu ÖZCAN YÜKSEK LİSANS TEZİ KENDİLENMİŞ MONOİK ATLANTİK SAKIZI POPULASYONUNDA GENETİK HARİTALAMA İÇİN POLİMORFİK YÖNTEM VE PRİMERLERİN BELİRLENMESİ BİYOTEKNOLOJİ
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ Burcu ÖZCAN YÜKSEK LİSANS TEZİ KENDİLENMİŞ MONOİK ATLANTİK SAKIZI POPULASYONUNDA GENETİK HARİTALAMA İÇİN POLİMORFİK YÖNTEM VE PRİMERLERİN BELİRLENMESİ BİYOTEKNOLOJİ
Karbapeneme Dirençli Acinetobacter baumannii Suşlarının PFGE Yöntemiyle Genotiplendirilmesi
 Karbapeneme Dirençli Acinetobacter baumannii Suşlarının PFGE Yöntemiyle Genotiplendirilmesi Yrd. Doç. Dr. Affan DENK Fırat Üniversitesi Tıp Fakültesi Enfeksiyon Hst. ve Klin. Mik. Araştırmacılar Yasemin
Karbapeneme Dirençli Acinetobacter baumannii Suşlarının PFGE Yöntemiyle Genotiplendirilmesi Yrd. Doç. Dr. Affan DENK Fırat Üniversitesi Tıp Fakültesi Enfeksiyon Hst. ve Klin. Mik. Araştırmacılar Yasemin
Giderilmesinde Farklı Uygulamalar
 Kavram Nedir? Kavramların Önemi Kavram Ö retimi Kavram Yanılgıları Kavram Ö retimi ve Yanılgılarının Giderilmesinde Farklı Uygulamalar Olgular Kavramlar lke ve Genellemeler Kuramlar ve Do a Kanunları Kavram
Kavram Nedir? Kavramların Önemi Kavram Ö retimi Kavram Yanılgıları Kavram Ö retimi ve Yanılgılarının Giderilmesinde Farklı Uygulamalar Olgular Kavramlar lke ve Genellemeler Kuramlar ve Do a Kanunları Kavram
Nesrin AKTEPE TANGU. ntangu@yalovabahce.gov.tr. Ege Üniversitesi Fen Bilimleri Enstitüsü Bahçe Bitkileri Anabilim Dalı 2012
 KİŞİSEL BİLGİLER Adı Soyadı Nesrin AKTEPE TANGU Unvan Mühendis (Dr.) Telefon 02268142520/1210 E-mail ntangu@yalovabahce.gov.tr Doğum Tarihi - Yeri 22.08.1970/Kalecik Fotoğraf Doktora Yüksek Lisans Lisans
KİŞİSEL BİLGİLER Adı Soyadı Nesrin AKTEPE TANGU Unvan Mühendis (Dr.) Telefon 02268142520/1210 E-mail ntangu@yalovabahce.gov.tr Doğum Tarihi - Yeri 22.08.1970/Kalecik Fotoğraf Doktora Yüksek Lisans Lisans
SANAL DĠLĠN DĠLĠMĠZDE YOL AÇTIĞI YOZLAġMA HAZIRLAYAN: CoĢkun ZIRAPLI Ġsmail ÇEVĠK. DANIġMAN: Faik GÖKALP
 SANAL DĠLĠN DĠLĠMĠZDE YOL AÇTIĞI YOZLAġMA HAZIRLAYAN: CoĢkun ZIRAPLI Ġsmail ÇEVĠK DANIġMAN: Faik GÖKALP SOSYOLOJĠ ALANI ORTAÖĞRETĠM ÖĞRENCĠLERĠ ARASI ARAġTIRMA PROJE YARIġMASI BURSA TÜRKĠYE BĠLĠMSEL VE
SANAL DĠLĠN DĠLĠMĠZDE YOL AÇTIĞI YOZLAġMA HAZIRLAYAN: CoĢkun ZIRAPLI Ġsmail ÇEVĠK DANIġMAN: Faik GÖKALP SOSYOLOJĠ ALANI ORTAÖĞRETĠM ÖĞRENCĠLERĠ ARASI ARAġTIRMA PROJE YARIġMASI BURSA TÜRKĠYE BĠLĠMSEL VE
2016 Ocak ENFLASYON RAKAMLARI 3 Şubat 2016
 2016 Ocak ENFLASYON RAKAMLARI 3 Şubat 2016 Ocak 2016 Tüketici Fiyat Endeksi ne(tüfe) ilişkin veriler Türkiye İstatistik Kurumu (TÜİK) tarafından 3 Şubat 2016 tarihinde yayımlandı. TÜİK tarafından aylık
2016 Ocak ENFLASYON RAKAMLARI 3 Şubat 2016 Ocak 2016 Tüketici Fiyat Endeksi ne(tüfe) ilişkin veriler Türkiye İstatistik Kurumu (TÜİK) tarafından 3 Şubat 2016 tarihinde yayımlandı. TÜİK tarafından aylık
ILIMAN İKLİM MEYVE TÜRLERİNDE ÇEŞİT VE TİPLERİN MOLEKÜLER TEKNİKLERLE KARAKTERİZASYONU
 ILIMAN İKLİM MEYVE TÜRLERİNDE ÇEŞİT VE TİPLERİN MOLEKÜLER TEKNİKLERLE KARAKTERİZASYONU Meryem YILDIZ ÖCAL Gıda Yüksek Mühendisi Manavgat Gıda Tarım ve Hayvancılık İlçe Müdürlüğü EĞİTİM BİLGİLERİ Eğitim
ILIMAN İKLİM MEYVE TÜRLERİNDE ÇEŞİT VE TİPLERİN MOLEKÜLER TEKNİKLERLE KARAKTERİZASYONU Meryem YILDIZ ÖCAL Gıda Yüksek Mühendisi Manavgat Gıda Tarım ve Hayvancılık İlçe Müdürlüğü EĞİTİM BİLGİLERİ Eğitim
ADIM ADIM YGS- LYS 92. ADIM KALITIM 18 GENETİK MÜHENDİSLİĞİ VE BİYOTEKNOLOJİ ÇALIŞMA ALANLARI
 ADIM ADIM YGS- LYS 92. ADIM KALITIM 18 GENETİK MÜHENDİSLİĞİ VE BİYOTEKNOLOJİ ÇALIŞMA ALANLARI GENETİK MÜHENDİSLİĞİ Belirli bir amaca yönelik olarak genetik madde üzerinde yapılan çalışmaları içerir. Canlıların
ADIM ADIM YGS- LYS 92. ADIM KALITIM 18 GENETİK MÜHENDİSLİĞİ VE BİYOTEKNOLOJİ ÇALIŞMA ALANLARI GENETİK MÜHENDİSLİĞİ Belirli bir amaca yönelik olarak genetik madde üzerinde yapılan çalışmaları içerir. Canlıların
Agaroz jel elektroforezi
 MOLEKÜLER TEKNİKLER Dr. Naşit İĞCİ Nevşehir Hacı Bektaş Veli Üniversitesi Moleküler Biyoloji ve Genetik Bölümü 4. Sınıf (2017-2018 Bahar) 2. NOT Agaroz jel elektroforezi PAGE daha çok proteinlerin ve küçük
MOLEKÜLER TEKNİKLER Dr. Naşit İĞCİ Nevşehir Hacı Bektaş Veli Üniversitesi Moleküler Biyoloji ve Genetik Bölümü 4. Sınıf (2017-2018 Bahar) 2. NOT Agaroz jel elektroforezi PAGE daha çok proteinlerin ve küçük
AKADEM SYENLER N ARA TIRMA YAPARKEN VE YAYINLATIRKEN KAR ILA TIKLARI GÜÇLÜKLER VE
 Uluslararası nsan Bilimleri Dergisi ISSN: 1303-5134 www.insanbilimleri.com 22.09.2004 AKADEM SYENLER N ARA TIRMA YAPARKEN VE YAYINLATIRKEN KAR ILA TIKLARI GÜÇLÜKLER VE BUNUN ÜZER NDE DOKTORA E T M N N
Uluslararası nsan Bilimleri Dergisi ISSN: 1303-5134 www.insanbilimleri.com 22.09.2004 AKADEM SYENLER N ARA TIRMA YAPARKEN VE YAYINLATIRKEN KAR ILA TIKLARI GÜÇLÜKLER VE BUNUN ÜZER NDE DOKTORA E T M N N
EGE ÜNĐVERSĐTESĐ FEN BĐLĐMLERĐ ENSTĐTÜSÜ (DOKTORA TEZĐ)
") EGE ÜNĐVERSĐTESĐ FEN BĐLĐMLERĐ ENSTĐTÜSÜ (DOKTORA TEZĐ) ASMA ( Vitis vinifera L. ) da GENOM HARĐTALAMASI: ÖNEMLĐ MORFOLOJĐK KARAKTERLERE VE FUNGAL KÖKENLĐ HASTALIKLARA YÖNELĐK AFLP ve SSR LĐNKAGE GRUPLARININ
EGE ÜNĐVERSĐTESĐ FEN BĐLĐMLERĐ ENSTĐTÜSÜ (DOKTORA TEZĐ) ASMA ( Vitis vinifera L. ) da GENOM HARĐTALAMASI: ÖNEMLĐ MORFOLOJĐK KARAKTERLERE VE FUNGAL KÖKENLĐ HASTALIKLARA YÖNELĐK AFLP ve SSR LĐNKAGE GRUPLARININ
Sait GEZGİN, Nesim DURSUN, Fatma GÖKMEN YILMAZ
 FARKLI ŞEKİL VE DOZLARDA UYGULANAN TKİ HÜMAS IN EKMEKLİK BUĞDAYIN BAYRAK YAPRAK BESİN ELEMENTİ KONSANTRASYONU, VERİM VE TANE PROTEİN KAPSAMINA ETKİLERİ Sait GEZGİN, Nesim DURSUN, Fatma GÖKMEN YILMAZ Bu
FARKLI ŞEKİL VE DOZLARDA UYGULANAN TKİ HÜMAS IN EKMEKLİK BUĞDAYIN BAYRAK YAPRAK BESİN ELEMENTİ KONSANTRASYONU, VERİM VE TANE PROTEİN KAPSAMINA ETKİLERİ Sait GEZGİN, Nesim DURSUN, Fatma GÖKMEN YILMAZ Bu
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTÜTÜSÜ
 ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTÜTÜSÜ DOKTORA TEZİ Muharrem YILMAZ BAZI FINDIK ÇEŞİT VE GENOTİPLERİNİN POMOLOJİK, MORFOLOJİK VE MOLEKÜLER KARAKTERİZASYONU BAHÇE BİTKİLERİ ANABİLİM DALI ADANA, 2009
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTÜTÜSÜ DOKTORA TEZİ Muharrem YILMAZ BAZI FINDIK ÇEŞİT VE GENOTİPLERİNİN POMOLOJİK, MORFOLOJİK VE MOLEKÜLER KARAKTERİZASYONU BAHÇE BİTKİLERİ ANABİLİM DALI ADANA, 2009
Siirt (Merkez) de Yetiştirilen Üzüm Çeşitlerinin Ampelografik Özelliklerinin Belirlenmesi Üzerine Bir Araştırma
 de Yetiştirilen Üzüm Çeşitlerinin Ampelografik Özelliklerinin Belirlenmesi Üzerine Bir Araştırma") Araştırma Makalesi / Research Article Iğdır Üni. Fen Bilimleri Enst. Der. / Iğdır Univ. J. Inst. Sci. & Tech. 1(3): 15-26, 2011 Iğdır Üniversitesi Fen Bilimleri Enstitüsü Dergisi Iğdır University Journal
Araştırma Makalesi / Research Article Iğdır Üni. Fen Bilimleri Enst. Der. / Iğdır Univ. J. Inst. Sci. & Tech. 1(3): 15-26, 2011 Iğdır Üniversitesi Fen Bilimleri Enstitüsü Dergisi Iğdır University Journal
Genetic Analysis of Grapevine Cultivars from the Eastern Mediterranean Region of Turkey, Based on SSR Markers
 TARIM BİLİMLERİ DERGİSİ 2009, 15(1) 1-8 ANKARA ÜNİVERSİTESİ ZİRAAT FAKÜLTESİ Genetic Analysis of Grapevine Cultivars from the Eastern Mediterranean Region of Turkey, Based on SSR Markers Serpil Gök TANGOLAR
TARIM BİLİMLERİ DERGİSİ 2009, 15(1) 1-8 ANKARA ÜNİVERSİTESİ ZİRAAT FAKÜLTESİ Genetic Analysis of Grapevine Cultivars from the Eastern Mediterranean Region of Turkey, Based on SSR Markers Serpil Gök TANGOLAR
ÖZGEÇMİŞ VE ESERLER LİSTESİ
 ÖZGEÇMİŞ VE ESERLER LİSTESİ ÖZGEÇMİŞ Adı Soyadı : Levent MERCAN Doğum Tarihi : 1 Ocak 1979 Öğrenim Durumu : Derece Bölüm/Program Üniversite Yıl Lisans Zootekni Ondokuz Mayıs Üniversitesi 2001 Y. Lisans
ÖZGEÇMİŞ VE ESERLER LİSTESİ ÖZGEÇMİŞ Adı Soyadı : Levent MERCAN Doğum Tarihi : 1 Ocak 1979 Öğrenim Durumu : Derece Bölüm/Program Üniversite Yıl Lisans Zootekni Ondokuz Mayıs Üniversitesi 2001 Y. Lisans
NOVACRETE Kİ MYASALİNİN ZEMİ N O ZELLİ KLERİ NE ETKİ Sİ Nİ N ARAŞTİRİLMASİNA İ Lİ ŞKİ N LABORATUVAR DENEYLERİ RAPORU
 YILDIZ TEKNİK ÜNİVERSİTESİ İNŞAAT FAKÜLTESİ - İNŞAAT MÜHENDİSLİĞİ BÖLÜMÜ NOVACRETE Kİ MYASALİNİN ZEMİ N O ZELLİ KLERİ NE ETKİ Sİ Nİ N ARAŞTİRİLMASİNA İ Lİ ŞKİ N LABORATUVAR DENEYLERİ RAPORU (25.06.2012)
YILDIZ TEKNİK ÜNİVERSİTESİ İNŞAAT FAKÜLTESİ - İNŞAAT MÜHENDİSLİĞİ BÖLÜMÜ NOVACRETE Kİ MYASALİNİN ZEMİ N O ZELLİ KLERİ NE ETKİ Sİ Nİ N ARAŞTİRİLMASİNA İ Lİ ŞKİ N LABORATUVAR DENEYLERİ RAPORU (25.06.2012)
EGE ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ (YÜKSEK LİSANS TEZİ)
") EGE ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ (YÜKSEK LİSANS TEZİ) ÖLÇEKLENEBİLİR H.264 VİDEO KODLAYICISI İÇİN SEVİYELENDİRİLEBİLİR GÜVENLİK SAĞLAYAN BİR VİDEO ŞİFRELEME ÇALIŞMASI Gül BOZTOK ALGIN Uluslararası
EGE ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ (YÜKSEK LİSANS TEZİ) ÖLÇEKLENEBİLİR H.264 VİDEO KODLAYICISI İÇİN SEVİYELENDİRİLEBİLİR GÜVENLİK SAĞLAYAN BİR VİDEO ŞİFRELEME ÇALIŞMASI Gül BOZTOK ALGIN Uluslararası
ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ
 ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ DİYARBAKIR İLİ ASMA GEN POTANSİYELİNİN RAPD (Random Amplified Polymorphic DNA) TEKNİĞİ İLE MOLEKÜLER ANALİZİ Hüseyin KARATAŞ Bahçe Bitkileri Ana
ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ DİYARBAKIR İLİ ASMA GEN POTANSİYELİNİN RAPD (Random Amplified Polymorphic DNA) TEKNİĞİ İLE MOLEKÜLER ANALİZİ Hüseyin KARATAŞ Bahçe Bitkileri Ana
Çizelge 1.1. İyonize radyasyon uygulamaları ile elde edilen mutant bitki türlerinin ülkelere göre dağılımı
 1. AMAÇ Mutasyonlar, kalıtım materyalinin (DNA ve RNA) fiziksel ve kimyasal yapısının değişmesi sonucu DNA dizininde, genetik açılım veya rekombinasyon kökenli olmayan, kalıtsal değişimler olarak tanımlanmaktadır
1. AMAÇ Mutasyonlar, kalıtım materyalinin (DNA ve RNA) fiziksel ve kimyasal yapısının değişmesi sonucu DNA dizininde, genetik açılım veya rekombinasyon kökenli olmayan, kalıtsal değişimler olarak tanımlanmaktadır
Mendel Genetiği, Kalıtım, Gen Mühendisliği ve Biyoteknoloji
 Mendel Genetiği, Kalıtım, Gen Mühendisliği ve Biyoteknoloji MENDEL GENETİĞİ Ebeveyn (ana-baba) ile oğul bireyler arasındaki benzerlik ve farklılıkların nasıl veya hangi oranlarda ortaya çıkabileceğini
Mendel Genetiği, Kalıtım, Gen Mühendisliği ve Biyoteknoloji MENDEL GENETİĞİ Ebeveyn (ana-baba) ile oğul bireyler arasındaki benzerlik ve farklılıkların nasıl veya hangi oranlarda ortaya çıkabileceğini
KARADENİZ BÖLGESİNDEN SEÇİLEN BAZI KIRMIZI AHUDUDU (Rubus ideaus L.) TİPLERİNİN GENETİK FARKLILIĞININ RAPD TEKNİĞİ İLE BELİRLENMESİ
 TİPLERİNİN GENETİK FARKLILIĞININ RAPD TEKNİĞİ İLE BELİRLENMESİ") AKDENİZ ÜNİVERSİTESİ ZİRAAT FAKÜLTESİ DERGİSİ, 2008, 21(2), 185 191 KARADENİZ BÖLGESİNDEN SEÇİLEN BAZI KIRMIZI AHUDUDU (Rubus ideaus L.) TİPLERİNİN GENETİK FARKLILIĞININ RAPD TEKNİĞİ İLE BELİRLENMESİ İlknur
AKDENİZ ÜNİVERSİTESİ ZİRAAT FAKÜLTESİ DERGİSİ, 2008, 21(2), 185 191 KARADENİZ BÖLGESİNDEN SEÇİLEN BAZI KIRMIZI AHUDUDU (Rubus ideaus L.) TİPLERİNİN GENETİK FARKLILIĞININ RAPD TEKNİĞİ İLE BELİRLENMESİ İlknur
Manisa da (Yuntdağı) Yetiştirilen Üzüm Çeşitlerinin Ampelografik Özelliklerinin Belirlenmesi Üzerine Araştırmalar 1
 Yetiştirilen Üzüm Çeşitlerinin Ampelografik Özelliklerinin Belirlenmesi Üzerine Araştırmalar 1") Ege Üniv. Ziraat Fak. Derg., 2006, 43(2):41-52 ISSN 1018-8851 Manisa da (Yuntdağı) Yetiştirilen Üzüm Çeşitlerinin Ampelografik Özelliklerinin Belirlenmesi Üzerine Araştırmalar 1 Harun ÇOBAN 2 Emrehan KÜEY
Ege Üniv. Ziraat Fak. Derg., 2006, 43(2):41-52 ISSN 1018-8851 Manisa da (Yuntdağı) Yetiştirilen Üzüm Çeşitlerinin Ampelografik Özelliklerinin Belirlenmesi Üzerine Araştırmalar 1 Harun ÇOBAN 2 Emrehan KÜEY
Hardy Weinberg Kanunu
 Hardy Weinberg Kanunu Neden populasyonlarla çalışıyoruz? Popülasyonları analiz edebilmenin ilk yolu, genleri sayabilmekten geçer. Bu sayım, çok basit bir matematiksel işleme dayanır: genleri sayıp, tüm
Hardy Weinberg Kanunu Neden populasyonlarla çalışıyoruz? Popülasyonları analiz edebilmenin ilk yolu, genleri sayabilmekten geçer. Bu sayım, çok basit bir matematiksel işleme dayanır: genleri sayıp, tüm
Sınıf ; Çalışma yaprağı 4
 Öğrencinin Adı ve soyadı ; Sınıf ; Çalışma yaprağı 4 F.8.2.2. Kalıtım Önerilen Süre: 10 ders saati Konu / Kavramlar: Gen, genotip, fenotip, saf döl, melez döl, baskın, çekinik, çaprazlama, cinsiyet, akraba
Öğrencinin Adı ve soyadı ; Sınıf ; Çalışma yaprağı 4 F.8.2.2. Kalıtım Önerilen Süre: 10 ders saati Konu / Kavramlar: Gen, genotip, fenotip, saf döl, melez döl, baskın, çekinik, çaprazlama, cinsiyet, akraba
MENDEL DIŞI KALITIM. Doç. Dr. Bengi ÇINAR KUL. Ankara Üniversitesi Veteriner Fakültesi Genetik Anabilim Dalı
 MENDEL DIŞI KALITIM Doç. Dr. Bengi ÇINAR KUL Ankara Üniversitesi Veteriner Fakültesi Genetik Anabilim Dalı Kalıtımla ilgili olarak Mendel kurallarına uymayan durumları içermektedir. Gen etkileşimleri Bağlı
MENDEL DIŞI KALITIM Doç. Dr. Bengi ÇINAR KUL Ankara Üniversitesi Veteriner Fakültesi Genetik Anabilim Dalı Kalıtımla ilgili olarak Mendel kurallarına uymayan durumları içermektedir. Gen etkileşimleri Bağlı
Ya! Satarım Bal Satarım Satı" Oyunu Etkinlik Ölçümü Sonuç Raporu. www.satisoyunu.com. Fuad Almeman Proje Ba" Danı"manı Aralık 2009
 U N I C O N D A N I! M A N L I K G R U B U Ya! Satarım Bal Satarım Satı" Oyunu Etkinlik Ölçümü Sonuç Raporu Fuad Almeman Proje Ba" Danı"manı Aralık 2009 www.satisoyunu.com Muallim Naci Cd. No. 53/2 Ortaköy
U N I C O N D A N I! M A N L I K G R U B U Ya! Satarım Bal Satarım Satı" Oyunu Etkinlik Ölçümü Sonuç Raporu Fuad Almeman Proje Ba" Danı"manı Aralık 2009 www.satisoyunu.com Muallim Naci Cd. No. 53/2 Ortaköy
DNA MİNİSATELLİT MARKIRLARINDAN YARARLANILARAK FİĞDE (Vicia sativa L.) TANE VERİMİNİN ÖNCEDEN BELİRLENMESİ OLANAKLARI
 TANE VERİMİNİN ÖNCEDEN BELİRLENMESİ OLANAKLARI") AKDENİZ ÜNİVERSİTESİ ZİRAAT FAKÜLTESİ DERGİSİ, 2005, 18(2), 169-174 DNA MİNİSATELLİT MARKIRLARINDAN YARARLANILARAK FİĞDE (Vicia sativa L.) TANE VERİMİNİN ÖNCEDEN BELİRLENMESİ OLANAKLARI Bilal AYDINOĞLU
AKDENİZ ÜNİVERSİTESİ ZİRAAT FAKÜLTESİ DERGİSİ, 2005, 18(2), 169-174 DNA MİNİSATELLİT MARKIRLARINDAN YARARLANILARAK FİĞDE (Vicia sativa L.) TANE VERİMİNİN ÖNCEDEN BELİRLENMESİ OLANAKLARI Bilal AYDINOĞLU
TARIMSAL YAPILAR VE SULAMA
 TARIMSAL YAPILAR VE SULAMA 4. YARIYIL KOD DERSLER İş Yükü AKTS K (saat) 0624401 Bahçe Bitkileri 150 5 3 0624402 Tarla Bitkileri 150 5 3 0624403 Bitki Koruma 150 5 3 0624404 Toprak Fiziği 120 4 3 0624405
TARIMSAL YAPILAR VE SULAMA 4. YARIYIL KOD DERSLER İş Yükü AKTS K (saat) 0624401 Bahçe Bitkileri 150 5 3 0624402 Tarla Bitkileri 150 5 3 0624403 Bitki Koruma 150 5 3 0624404 Toprak Fiziği 120 4 3 0624405
1 OCAK 31 ARALIK 2009 ARASI ODAMIZ FUAR TEŞVİKLERİNİN ANALİZİ
 1 OCAK 31 ARALIK 2009 ARASI ODAMIZ FUAR TEŞVİKLERİNİN ANALİZİ 1. GİRİŞ Odamızca, 2009 yılında 63 fuara katılan 435 üyemize 423 bin TL yurtiçi fuar teşviki ödenmiştir. Ödenen teşvik rakamı, 2008 yılına
1 OCAK 31 ARALIK 2009 ARASI ODAMIZ FUAR TEŞVİKLERİNİN ANALİZİ 1. GİRİŞ Odamızca, 2009 yılında 63 fuara katılan 435 üyemize 423 bin TL yurtiçi fuar teşviki ödenmiştir. Ödenen teşvik rakamı, 2008 yılına
Yrd.Doç.Dr. Yosun MATER
 * Yrd.Doç.Dr.Yosun MATER Yrd.Doç.Dr. Yosun MATER *Bitki nüklear, mitokondriyal ve kloroplast DNA'ları *Burada yer alan bugünkü bilgilerimizin çoğu, moleküler evrim mekanizması ve oranları kullanılarak
* Yrd.Doç.Dr.Yosun MATER Yrd.Doç.Dr. Yosun MATER *Bitki nüklear, mitokondriyal ve kloroplast DNA'ları *Burada yer alan bugünkü bilgilerimizin çoğu, moleküler evrim mekanizması ve oranları kullanılarak
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
 ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ YEREL MISIR POPULASYONLARININ MORFOLOJİK VE DNA MOLEKÜLER İŞARETLEYİCİLERİNDEN SSR TEKNİĞİ İLE KARAKTERİZASYONU TARLA BİTKİLERİ ANA BİLİM DALI
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ YEREL MISIR POPULASYONLARININ MORFOLOJİK VE DNA MOLEKÜLER İŞARETLEYİCİLERİNDEN SSR TEKNİĞİ İLE KARAKTERİZASYONU TARLA BİTKİLERİ ANA BİLİM DALI
TÜRKĠYE ve DÜNYA BAĞCILIĞI. Dr. Selçuk KARABAT 1
 TÜRKĠYE ve DÜNYA BAĞCILIĞI Dr. Selçuk KARABAT 1 Bağcılığın Tarihi Dünya da çok geniş bir alanda yapılmakta olan bağcılıkta asma veya omca (Vitis sp.) denilen bitki yetiştirilmektedir. Milyonlarca yıl öncesine
TÜRKĠYE ve DÜNYA BAĞCILIĞI Dr. Selçuk KARABAT 1 Bağcılığın Tarihi Dünya da çok geniş bir alanda yapılmakta olan bağcılıkta asma veya omca (Vitis sp.) denilen bitki yetiştirilmektedir. Milyonlarca yıl öncesine