POLİMERLERİN LEISHMANIA- KONAK HÜCRE ETKİLEŞİMİ ÜZERİNE ETKİSİNİN İNCELENMESİ
|
|
|
- Berk Tarhan
- 6 yıl önce
- İzleme sayısı:
Transkript
1 YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ POLİMERLERİN LEISHMANIA- KONAK HÜCRE ETKİLEŞİMİ ÜZERİNE ETKİSİNİN İNCELENMESİ Biyomühendis Serhat ELÇİÇEK FBE Biyomühendislik Anabilim Dalından Hazırlanan YÜKSEK LİSANS TEZİ Tez Danışmanı: Yrd. Doç. Dr. Melahat BAĞIROVA (YTÜ) İSTANBUL, 2009
2 İÇİNDEKİLER Sayfa SİMGE LİSTESİ v KISALTMA LİSTESİ vi ŞEKİL LİSTESİ. vii ÇİZELGE LİSTESİ. xi ÖNSÖZ ÖZET.. xii..xiii ABSTRACT... xiv 1. GİRİŞ GENEL BİLGİLER Leishmania Parazitleri Parazitlerin Morfolojisi Parazitlerin Hayat Döngüsü Leishmaniasis Kutanöz Leishmaniasis Mukokutanöz Leishmaniasis Visseral Leishmaniasis İmmunoloji Leishmania Parazitlerinin Konak Hücre İçindeki Yaşamı Leishmania Parazitlerinin Kültürünün Yapılması Kullanılan Besiyerleri Mononükleer Fagositik Sistem Hücrelerinin Gelişimi J774 Hücre Hattı Makrofajların Aktivasyonu ve Fonksiyonları Fagositoz Salgılama Antijen Sunma Makrofajların Özellikleri Nitrik oksit (NO) Polimerler Polimerlerin Genel Özellikleri ve Sınıflandırılmaları Polielektrolitler Homopolielektrolitler Poliamfolitler Polimerlerin Tıp ve Biyoteknolojide Kullanımı Poliakrilik Asit Biyouyumluluk ve Toksisite Testleri ii
3 (4,5-dimetiltiazol-2-il)-2,5-difeniltetrazolyum (MTT) Testi DENEYSEL ÇALIŞMALAR Deneysel Çalışmalarda Kullanılan Cihazlar ve Kimyasal Maddeler Deneysel Çalışmalarda Kullanılan Cihazlar Deneysel Çalışmalarda Kullanılan Kimyasal Maddeler ve Besiyerleri Çözeltilerin Hazırlanması x PBS Tampon Hazırlanması x Tripsin solusyonu RPMI 1640 Medyumunun Hazırlanması Hücre Dondurma Medyumunun Hazırlanması MTT Solüsyonunun Hazırlanması MTT Reaksiyon Sonlandırıcı Solüsyonun Hazırlanması Poliakrilik Asit (50 mg/ml, MA: 100,000 g/mol) Çözeltisinin Hazırlanması Poliakrilik Asit (50 mg/ml, MA: 30,000) Çözeltisinin Hazırlanması Bağlayıcı Tampon (Binding Buffer) Çözeltisi Hazırlaması Annexin V-FITC Solüsyonu Hazırlaması AAD Solüsyonu Hazırlanması Hücre Kültürü Teknikleri Hücrelerin Kriyobanktan Çıkarılması J774 Hücre Hattının Kriyobanktan Çıkarılması MONI/EP126 L. infantum Suşlarının Kriyobanktan Çıkarılması Hücre Hatlarının Kültürü J774 Hücre Hattının Kültürü Tripan Mavisi ile J774 Hücre Canlılığının Tayin Edilmesi ve Hücre Sayımı MONI/EP126 Leishmania Promastigot Kültürünün Yapılması Parazitlerin Thoma Lamında Sayımı Hücrelerin Dondurulması ve Saklanması J774 Hücrelerinin Dondurulması ve Saklanması MONI/EP126 Promastigotlarının Dondurulması ve Saklanması Polimerlerin Toksik Etkisinin MTT Yöntemi ile İncelenmesi PAA nın J774 Hücrelerindeki 48 Saatlik Toksik Etkisinin MTT Yöntemi ile İncelenmesi PAA nın Ep126 Promastigotlarına 48 Saatlik Toksik Etkisinin MTT Yöntemi ile İncelenmesi J774 Makrofaj Hücrelerinin Annexin-V&7-AAD İşaretlenmesi Promastigotların Giemsa ile Boyanması Makrofaj Hücrelerinin L. infantum ile Enfekte Edilmesi Enfekte Olmuş Makrofajların Giemsa ile Boyanması Enfektifliğin Hesaplanması İstatistik Analiz DENEYSEL SONUÇLAR VE TARTIŞMALAR Poliakrilik Asitin J774 Hücrelerine Apoptotik Etkisinin İncelenmesi PAA nın J774 Hücre Kültüründe Toksik Etkisinin İncelenmesi Molekül Ağırlığı Olan PAA nın J774 Hücreleri Üzerindeki Toksik Etkisinin İncelenmesi Molekül Ağırlığı Olan PAA nın J774 Hücreleri Üzerindeki Toksik Etkisinin İncelenmesi PAA nın Leishmania promastigotlarına Toksik Etkisinin İncelenmesi iii
4 6.3.1 Molekül Ağırlığı 100,000 Olan PAA nın Ep126 Promastigotları Üzerindeki Toksik Etkisinin İncelenmesi Molekül Ağırlığı 30,000 Olan PAA nın Ep126 Promastigotları Üzerindeki Toksik Etkisinin İncelenmesi PAA ya Maruz Kalmış Ep126 Promastigotlarının J774 Makrofaj Hücreleri ile Etkileşimi ve Enfektifliğinin İncelenmesi Poliakrilik Asite (MA:100,000) Maruz Kalmış Ep126 Promastigotlarının J774 Hücreleri ile Etkileşimi ve Enfektifliğinin İncelenmesi PAA ya (15 mg/ml, MA:100,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi PAA ya (5 mg/ml, MA:100,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi PAA ya (1 mg/ml, MA:100,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi Poliakrilik Asite (MA:30,000) Maruz Kalmış Ep126 Promastigotlarının J774 Hücreleri ile Etkileşimi ve Enfektifliğinin İncelenmesi PAA ya (15 mg/ml, MA:30,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi PAA ya (5 mg/ml, MA:30,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi PAA ya (1 mg/ml, MA:30,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi Tartışma KAYNAKLAR İNTERNET KAYNAKLARI ÖZGEÇMİŞ 100 iv
5 SİMGE LİSTESİ Da g Dalton Gram µg Mikrogram μl μm mg ml mm MA Mikrolitre Mikrometre Miligram Mililitre Milimolar Moleküler ağırlığı ºC Santigrat derece v
6 KISALTMA LİSTESİ AIDS Edinilmiş bağışık yetersizliği hastalığı DMSO Dimetil sülfoksit DNA Deoksiribo nükleik asit EP126 Leishmania promastigot kültürü ETDA Etilendiamin tetraasetik asit FBS Fetal sığır serumu FCS Fetal calf serum FDA Amerikan Gıda ve İlaç Dairesi FDS Fetal dana serum HCl Hidrojen klorür HEPA Yüksek etkili partikül yakalayıcı HEPES N-2-hidroksi etilpiperazin N-2 etansulfonik asit HPMA N-(2-hidroksipropil) metakrilamid KCl Potasyum klorür KH 2 PO 4 Potasyum dihidrojen fosfat MTS 3-(4,5-dimetiltiazol-2-il-)-5-(3-karboksimetilmethoksifenil)-2-(-4-sulfofenil)-2H-tetrazolyum MTT 3-(4,5-dimetiltriazol-2-il)-2,5- difeniltetrazolium bromid NaCl Sodyum klorür Na 2 HPO 4 Di sodyum hidrojen fosfat NIPAA N-izopropil akril amid NNN Novy-MacNeal ve Nicole medyum PAA Poliakrilik asit PBS Fosfat tampon solüsyonu PEG Polietilen glikol RNA Ribonükleik asit SDS Sodyum dodesil sülfat UV Ultraviyole vi
7 ŞEKİL LİSTESİ Sayfa Şekil 2.1 Leishmania parazitlerinin formları... 5 Şekil 2.2 Leishmania parazitlerinin hayat döngüsü... 6 Şekil 3.1 Makrofaj aktivasyonu Şekil 4.1 Polielektrolitlerin sınıflandırılması (a)polikatyon (b)polianyon (c)poliamfolit.23 Şekil 4.2 Poliakrilik asitin sulu çözeltideki dissosiyasyonu Şekil 4.3 Polimer-ilaç, polimer-protein ve polimer-dna konjugat sistemleri Şekil 4.4 Poliakrilik asitin yapısı Şekil 6.1 PAA bulunmayan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) Şekil 6.2 PAA (2,5 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) Şekil 6.3 PAA (2,5 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) Şekil 6.4 PAA (250 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) Şekil 6.5 PAA (250 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) Şekil 6.6 PAA (2500 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) Şekil 6.7 PAA (2500 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) Şekil 6.8 J774 hücrelerinin 48 saat inkübasyon sonrası morfolojik görüntüsü (MTT eklenmeden önce) (20x) Şekil 6.9 J774 hücrelerinde formazan kristallerinin oluşumu (48 saat inkübasyon sonrası, MTT eklendikten sonra) (20x) Şekil 6.10 PAA nın 2500 µg/ml konsantrasyonunun J774 hücreleri üzerindeki sitopatik etkisi (20x) Şekil 6.11 PAA nın 500 µg/ml konsantrasyonunun J774 hücreleri üzerindeki sitopatik etkisi (20x) Şekil 6.12 PAA nın 250 µg/ml konsantrasyonunun J774 hücreleri üzerindeki sitopatik etkisi (20x) Şekil 6.13 PAA nın 2,5 µg/ml konsantrasyonunun J774 hücreleri üzerindeki sitopatik etkisi (20x) Şekil 6.14 PAA nın 2500 µg/ml konsantrasyonun J774 hücreleri üzerindeki sitopatik etkisi (20x) Şekil 6.15 PAA nın 1000 µg/ml konsantrasyonun J774 hücreleri üzerindeki sitopatik etkisi (20x) Şekil 6.16 PAA nın 500 µg/ml konsantrasyonun J744 hücreleri üzerindeki sitopatik etkisi (20x) Şekil 6.17 PAA nın 2,5 µg/ml konsantrasyonun J774 hücreleri üzerindeki sitopatik etkisi (20x) Şekil 6.18 L.infantum promastigotlarının morfolojik görüntüsü (Giemsa boyama, 100x) vii
8 Şekil 6.19 Şekil 6.20 Şekil 6.21 Şekil 6.22 Şekil 6.23 Şekil 6.24 Şekil 6.25 Şekil 6.26 Şekil 6.27 Şekil 6.28 Şekil 6.29 Şekil 6.30 Şekil 6.31 Şekil 6.32 Şekil 6.33 Şekil 6.34 Şekil 6.35 Şekil 6.36 Şekil 6.37 Şekil 6.38 Şekil 6.39 Şekil 6.40 Şekil 6.41 Şekil 6.42 Şekil 6.43 (48 saat inkübasyon sonrası) mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).72 viii
9 Şekil 6.44 Şekil 6.45 Şekil 6.46 Şekil 6.47 Şekil 6.48 Şekil 6.49 Şekil 6.50 Şekil 6.51 Şekil 6.52 Şekil 6.53 Şekil 6.54 Şekil 6.55 Şekil 6.56 Şekil 6.57 Şekil 6.58 Şekil 6.59 Şekil 6.60 Şekil 6.61 Şekil 6.62 Şekil 6.63 Şekil 6.64 Şekil 6.65 Şekil 6.66 Şekil 6.67 Şekil 6.68 Şekil mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).73 5 mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).73 5 mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).74 5 mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).74 5 mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).75 5 mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).75 1 mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).76 1 mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).76 1 mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).77 1 mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).77 1 mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).78 1 mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).82 5 mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).83 5 mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).83 5 mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).84 5 mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).85 5 mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).86 1 mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile ix
10 Şekil 6.70 Şekil 6.71 Şekil 6.72 Şekil 6.73 enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x).86 1 mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).87 1 mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x).88 1 mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) x
11 ÇİZELGE LİSTESİ Sayfa Çizelge 2.1 Leishmania parazitlerinin sınıflandırılması... 4 Çizelge 2.2 Leishmaniasis kliniği ve etkenlerine göre sınıflandırılması... 7 Çizelge 4.1 Bazı biyomedikal polimerlerin önemli kullanım alanları Çizelge 4.2 Akrilik asitin bazı özellikleri Çizelge 6.1 PAA nın (MA:100,000 ) farklı konsantrasyonlarının J774 hücrelerinin canlılığına etkisi Çizelge 6.2 PAA nın (MA:30,000) farklı konsantrasyonlarının J774 hücrelerinin canlılığına etkisi Çizelge 6.3 MA:100,000 olan PAA nın Ep126 promastigotlarının canlılığına etkisi Çizelge 6.4 MA:100,000 olan PAA nın Ep126 promastigotlarına in vitro etkisi Çizelge 6.5 MA:30,000 olan PAA nın Ep126 promastigotlarına in vitro etkisi Çizelge 6.6 Poliakrilik Asite (MA:100,000) Maruz Kalmış Ep126 Promastigotlarının J774 Hücrelerine Enfektifliği Çizelge 6.7 Poliakrilik Asite (MA:30,000) Maruz Kalmış Ep126 Promastigotlarının J774 Hücrelerine Enfektifliği xi
12 ÖNSÖZ Lisansüstü eğitimim boyunca akademik bilgisini ve tecrübesini benden esirgemeyen çok değerli danışman Hocam Yrd. Doç. Dr. Melahat BAĞIROVA'ya, çalışmalarım boyunca aynı ilgi ve desteği gösteren değerli Hocam Prof. Dr. Adil M. ALLAHVERDİYEV'e, verdiği mali destekten dolayı TÜBİTAK ve ek destek sağlayan Develet Planlama Teşkilatı na (DPT) teşekkür ederiz. Projeye ek destek sağlayan DPT projesinin ilk yürütücüsü Prof. Dr. Mehmet Mustafa AKDESTE ye teşekkürü bir borç bilip yakın zamanda kaybettiğimiz hocamızı saygı ile anıyoruz. Sözkonusu DPT projesini devralan ve büyük bir özveri ile yürütmekte olan Kimya- Metalurji Fakültesi Dekanı Prof. Dr. Sabriye PİŞKİN e de yine teşekkürü borç biliriz. Ayrıca bu çalışmanın yerine yetirimesine imkan sağlayan Kimya-Metalurji Fakültesi Dekanlığına, Fen Bilimeri Enstitüsüne, Biyomühendilik Bölüm Başkanlığına ve bütün değerli Hocalarıma teşekkür ederim. Deneysel çalışmam boyunca benden yardımlarını esirgemeyen arkadaşlarım Sezen CANIM, Rabia ÇAKIR, Melike ERSÖZ, Olga Nehir ÖZTEL, Serap YEŞİLKIR ve Emrah Şefik ABAMOR a teşekkür ederim. Bu tez çalışması SBAG-4007 nolu TÜBİTAK projesi tarafından desteklenmiştir. xii
13 ÖZET Leishmaniasis Dünyanın ve Türkiye nin ciddi halk sağlığı problemlerinden biridir. Hastalığın tedavisinde kullanılan ilaçlar yetersiz olmakla birlikte pahalı ve toksiktir. Aşı geliştirilmesi için çeşitli yöntemler denenmesine rağmen şimdiye kadar etkin bir aşı geliştirilememiştir. Son yıllarda polielektrolitlerin, hücre ve biyomakromoleküllerle in vitro ve in vivo etkileşimi üzerine yapılan çalışmalar giderek artmaktadır. Özellikle in vivo yapılan bazı çalışmalarda polielektrolitlerin immun cevabı artırdığı ve adjuvant etkisinin olduğu gösterilmiştir. Bu çalışmada ilk kez olarak Poliakrilik asitin (PAA) farklı molekül ağırlıklarının in vitro kültür ortamında Leishmania promastigotları ile konak hücre (makrofaj) arasındaki etkileşimine etkisi incelenmiştir. Deneylerde Visseral leishmaniasis (VL) etkeni olan L.infantum un promastigot kültürü (MONI/EP126), J774 makrofaj konak hücre kültürü kullanılmıştır. PAA nın ise, molekül ağırlığı 30,000 ve 100,000 olan farklı konsantrasyonları incelenmiştir. PAA nın hücreler üzerine etkisi flow sitometri (apoptotik- nekrotik), MTT ve tripan mavisi (hücre canlılığı ve IC 50 ), mikroskobik (hücre sayımı, % enfektiflik ve morfoloji) gibi çeşitli yöntemler incelendi. Veriler, SPSS version 16.0 for Windows programı ile yapıldı ve p<0,05 değeri istatistiki olarak anlamlı kabul edildi. Sonuçlar, PAA nın incelenen 1 mg/ml konsantrasyonunda promastigotları, 100 µg/ml onsantrasyonlarda ise makrofajlara toksik etki etmediğini, ancak bunun üzerinde olan konsantrasyonlarda ise hücrelerin proliferasyonunu inhibe ettiğini gösterdi. Ayrıca PAA nın incelenen yaklaşık tüm konsantrasyonlarında, parazitlerin enfektifliğinin önemli ölçüde azaldığı gözlendi (p<0,05). Elde edilen sonuçlar Leishmania da immunojen olan bazı moleküllerle, polimerlere dayalı konjugat oluşturulması için PAA nın önemli adaylardan biri olduğunu ortaya koydu. Anahtar kelimeler: PAA, J774, L.infantum, makrofaj, toksisite, MTT, hücre kültürü, amastigot xiii
14 ABSTRACT In Turkey and around the world, Leishmaniasis is considered as one of the very serious health issues. Drugs that are used for the treatment of this disease are not only inadequate but also expensive and toxic. Although many approaches for the vaccine development are essayed, an effective vaccine could not been developed yet. Recently, studies on in vitro and in vivo interactions between polyelectrolites and both cells and biomacromolecules are increasing. Especially, some in vivo studies have shown that polyelectrolites induced immune response and also exerted an adjuvant effect. In this study, for a first time, poliacrilic acid (PAA) of different moleculer weights was essayed for their effects on the in vitro interaction of Leishmania promastigots and host cell (macrophage). Visseral leishmaniasis (VL) causing promastigot culture of L.infantum (MONI/EP126) and J774 macrophage host cell culture are used during this study. Furthermore, different concentrations of PAA that have molecular weights of 30,000 and 100,000 are studied. Effects of PAA on cells were monitored using a variety of methods like flow cytometry (apoptic-necrotic), MTT and trypan blue (cell viability and IC 50), microscopy (cell count, % infectivity and morphology). All the data were processed using SPSS version 16.0 for Windows program and p<0,05 value is accepted statistically meaningful. Results have revealed that PAA of 1 mg/ml concentration did not show a toxic effect on promastigots, while PAA of 100 µg/ml concentration did not show a toxic effect on macrophages. However, in case of higher concentrations, PAA was shown to inhibit cell proliferation. Furthermore, for almost all essayed concentrations of PAA, parasite infectivity was observed to be significantly decreased (p<0,05). This data obtained revealed out that PAA is an important candidate for the construction of polymer based conjugates with some molecules that are immunogenic in Leishmania. Keywords: PAA, J774, L.infantum, macrophage, toxcicity, MTT, cell culture, amastigot xiv
15 1 1. GİRİŞ Dünyada 12 ile 40 milyon arasında kişinin Leishmania ile enfekte olduğu, 350 milyona yakın kişinin ise bu hastalığa yakalanma riski altında olduğu bilinmektedir. Her yıl 1,0-1,5 milyon insan deri leishmaniasisine, kişi ise visseral leishmaniasis e yakalanmaktadır (Desjeux, 2001). Türkiyede ise yaklaşık 20 milyon kişi bu enfeksiyonun tehditi altındadır. Ülkenin değişik bölgelerinde visseral ve özellikle de kutanöz formlarına rastlanmaktadır. Her yıl yeni olgu sayısı da yaklaşık 5000 kişi civarındadır (Özcel, 1995). Hastalığın tedavisinde kullanılan ilaçlar oldukça toksikdir. Son yıllarda hastalık etkenlerinde kullanılan ilaçlara, vektörlerinde ise insektisitlere karşı dirençli olduğu bilinmektedir. Ayrıca birçok çalışma yapılmasına rağmen leishmaniaya karşı halen etkin bir aşı geliştirilememiştir. Leishmaniasisin Türkiye nin ciddi halk sağlığı problemi olmasına rağmen ülkemizde bu hastalığa karşı aşı geliştirilmesi ile ilgili çalışma bulunmamaktadır. Son yıllarda polielektrolitlerin, hücre ve biyomakromoleküllerle in vitro ve in vivo etkileşimi üzerine yapılan çalışmalar giderek artmaktadır. Özellikle yapılan bazı çalışmalarda polielektrolitlerin (Poliakrilik asit, Poli-4-vinilpiridin ve Poli-2-metil-5-vinilpiridin gibi) in vivo ve in vitro koşullarda immun cevabı artırdığı ve adjuvant etkisi olduğu belirlenmiştir. Ancak, adjuvant özelliği olan polielektrolitlerin parazitik enfeksiyon ajanıyla etkileşimi sonucunda in vitro ve in vivo koşularda oluşacak immun yanıt ve enfektiflik konusunda hiçbir çalışma bulunmamaktadır. Parazit ile konak arasındaki etkileşimlerin temelini parazit yüzeyinde bulunan antijenik özellikler gösteren ligandlar (LPG, Gp63 gibi) ile konak yüzeyinde bulunan reseptörler oluşturur. Bu nedenle polielektrolitlerin in vitro parazit konak etkileşimine etkisinin incelenmesinde, konak ve parazit yüzey yapılarının varlığı önemli olabilir. Ancak polimerlerin, LPG ve diğer ligandlar ile oluşturacağı konjugatların in vivo modellerde incelenmesinden önce, in vitro parazit ve konak hücre etkileşimine etkisinin incelenmesi sonucunda elde edilen bilgiler ileride oluşturulacak konjugatın ne kadar değerli olabilmesi açısından önemlidir. Bu polimerlerden biri de zayıf anyonik polielektrolit olan PAA olup daha önce yapılan çalışmalarda önemli ölçüde adjuvant etkisi olduğu gösterilmiştir. Buna göre de bu çalışmada ilk kez olarak Poliakrilik asitin farklı molekül ağırlıklarının in vitro kültür ortamında Leishmania promastigotları ile konak hücre (makrofaj) arasındaki etkileşimini incelenmiştir. Bu amaçla; 1. PAA nın farklı molekül ağırlıkları ve konsantrasyonlarının promastigotlar üzerine in vitro etkisi incelendi.
16 2 2. PAA nın farklı molekül ağırlıkları ve konsantrasyonlarının makrofajlar üzerine in vitro etkisi incelendi. 3. Promastigotlar için toksik ve toksik olmayan konsantrasyonlar belirlendikten sonra parazitlerin % enfektiflik derecesi belirlendi
17 3 2. GENEL BİLGİLER 2.1 Leishmania Parazitleri Leishmania parazitlerinin neden olduğu Leishmania hastalığı (Leishmaniasis), klinik ve epidemiyolojik açıdan çok çeşitlilik gösteren ve ciddi bir halk sağlığı problemi olarak tüm dünyada önemini koruyan bir hastalıktır (Cattand, 2006). Zorunlu hücre içi parazit olan Leishmania, trypanozom ailesinin kinetoplastida sınıfına aittir. Leishmania parazitleri ile enfekte dişi tatarcıkların kan emmesi sırasında hastalık bulaştırır (Ak vd., 1995; Desjeux, 2004). Leishmania parazitleri ilk kez olarak Leishman tarafından 1900 yılında Hindistan'da dizanteriye yakalanan bir hastanın dalağında görülmüştür. Daha sonra Donovan Madras'ta viseral leishmaniasisli hastaların dalağından elde ettiği örneklerde paraziti görmüş ve bulgularını 1903 yılında yayınlamıştır. Ross ise sonraki yıllarda paraziti L. donovani olarak adlandırmıştır (Töre, 1996; Özçelik, 1999). Leishmania parazitlerinin in vitro kültürü ilk kez Nicolle tarafından 1908 de yapılmış, tatarcıklardaki evrimi ise sonraki yıllarda çeşitli araştırıcılar tarafından açıklığa belirlenmiştir. Nicolle ve Compte köpeklerde Leishmania parazitlerini tespit edilmiştir. Nicole tarafından 1908 yılında bu parazite L. infantum adı verilmiştir (Özcel, 1995). Leishmania cinsi içinde farklı türler ve alt türler bulunur (Saygı, 2002). Işık mikroskobunda bütün türler morfolojik olarak aynı görünüme sahip olduğundan; neden oldukları hastalıklar, epidemiyolojik, coğrafik dağılım, serolojik, immunolojik, biyokimyasal ve biyolojik özelliklerine dayanarak sınıflandırma yapılmıştır. Son yıllarda sınıflandırmada izoenzimlerin araştırılması, monoklonal antikorlar ya da parazitin DNA yapısının incelenmesi de dikkate alınmıştır. Buna rağmen sınıflandırma da sorunlar devam etmektedir. Dünya Sağlık Örgütü'nün onayladığı sınıflandırma Çizelge 2.l de verilmiştir (Altıntaş, 1995; Çeliksöz, 1997).
18 4 Çizelge 2.l Leishmania parazitlerinin sınıflandırılması 2.2 Parazitlerin Morfolojisi Leishmania parazitleri insan ve diğer memelilerde amastigot, vektörlerde ise (sand fly, tatarcık, yakarca) promastigot olmak üzere iki farklı morfolojik şekilde bulunur. Parazitlerin amastigot formu, 1 3 µm eninde, 2-5 µm boyunda, oval ve hareketsizdir. Sitoplazmada arka uca yakın büyük bir nukleus ve nukleusa yakın kinetoplast bulunur (Şekil 2.1). Kinetoplast elektron mikroskobunda disk biçiminde görülür ve boyutu 0,4 0,8 µm arasında değişir. Giemsa ile boyanan simirlerde amastigotun sitoplazması soluk mavi, çekirdeği ise koyu kırmızı renkte görülür. Kinetoplast çekirdeğin yanında parlak kırmızı veya mor renkte boyanır. Kinetoplast'ta çekirdekten farklı bir DNA bulunur. Bu DNA'nın dizilimi türlere göre farklılık gösterdiği için tür belirlenmesinde kullanılır. Kinetoplastın genetik bir devamlılığı olduğu ve çekirdekten önce bölündüğü belirlenmiştir. Kinetoplast ta koyu boyanan parabazal cisim ve yanında nokta şeklindeki blefaroplasttan çıkıp ön kısımda sonlanan bir aksonem vardır. Elektron mikroskobu ile yapılan gözlemlerde parazit plazma membranının tipik olarak üç katlı olduğu, hücreye destek görevi de yapan 2-4 nm'lik bir yapı olan pelikül altı mikrotübülleri bulunmaktadır (Unat vd., 1995). Makromoleküllerin hücre içine alınışında önemli rol oynayan bu mikrotübüllerin sayıları ve merkezden uzaklıkları Leishmania tür ayrımında

19 5 kullanılmaktadır. Bütün Leishmania türlerinde sitoplazmada tek bir mitokondri bulunmaktadır. Lizozomların, suda erimeyen polifosfat asidinin, peroksizomların ve RNA içeren volutin taneciklerinin, parazitin beslenmesinden ve salgı kontrolünden sorumlu olduğu belirlenmiştir (Orhan ve Yaşarol, 1981). Amastigotlar, kamçı cebi içinde aktif ve serbest olmayan bir kamçıya sahiptir. Tatarcıkların bağırsaklarında ve in vitro kültür ortamlarında bulunan ve parazitin hücre dışı formu olan promastigotların ise, µm boyunda, 1,5-3 µm genişlikte ve ön uçtan çıkan serbest bir kamçısı bulunmaktadır (Şekil 2.1) (Garcia, 2001). Kamçının bir çift merkez ve dokuz çift periferik fibril çiftinden oluşan bir aksonemi vardır. Ön uçta yuvarlak veya at nalı şeklinde kinetoplast vardır. Kinetoplastın ön ve kamçının dip kısmında ise bleforablast bulunur. Çekirdek ve çekirdekçik merkezde bulunur ve çekirdek zarında porlar bulunur. Ayrıca sitoplazma içinde golgi aygıtı ve endoplazmik retikulum gibi organeller vardır (Bryceson, 1996). Şekil 2.1 Leishmania parazitlerinin formları (Chiodini vd., 2003) 2.3 Parazitlerin Hayat Döngüsü Dişi tatarcıklar enfekte konaktan kan emerken amastigotlar ile enfekte olurlar. Kan ile alınan amastigotların bir kısmı sindirilirken, bir kısmı da orta bağırsakta bölündükten sonra kamçı oluşturur ve metasiklik (enfektif) promastigot formuna dönüşürler. Peritrofik membran içinde çoğalarak, membranı eriten promastigotlar, torasik mideye geçerek kemotaksis ile özafagus ve farinkse tutunurlar. Özafagustan ayrılan daha küçük vektörün formlar 7. gününde ağız kısımlarında ve hortumda görülebilir. Ağız kısmında bulunan enfektif promastigotlar, tatarcıkların kan emmesini düzenleyen kardiak kapağa zarar vererek, kan akımının tersi yönünde tatarcığın vücudundan omurgalı konağa geçerler. Makrofajların promastigotlarla 4 8 saat içinde enfekte olduğu ve 24 saat sonra amastigotların çoğaldığı belirlenmiştir (Garcia, 2001). Konağa geçen promastigotların

20 6 bir kısmı, kan dolaşımına katılarak konak tarafından öldürülür. Ancak geri kalan promastigotlar deri makrofajları veya dendritik hücreler tarafından reseptör temelli fagositoz ile fagosite edilir. Fagosite edilen promastgotlar, amastigot formuna dönüşerek çoğalırlar ve hücreyi patlatarak serbest kalırlar. Hücreyi parçalayarak serbest hale geçen bu amastigotlar, yeniden makrofajları enfekte ederek dalak, karaciğer ve kemik iliği gibi retikuloendotelial sistem organlarına yerleşirler (Bryceson 1996; Jeronimo ve Pearson, 1992). Enfekte olmuş konakların kan ve lezyonlarında hem serbest amastigotlar, hem de enfekte makrofajlar vardır. Tatarcık, enfekte konaktan tekrar kan emdiğinde, hem serbest amastigotları, hem de enfekte makrofajları alır. Amastigotlar tekrar tatarcığın mide kanalında gelişim sürecini geçirirerek, promastigot forma dönüşürler. Bu döngü tatarcığın beslenmek için bir başka konak bulmasıyla devam etmekte ve tüm bu döngü özellikle çevre ısısına bağlı olarak 4 25 gün sürmektedir (Despommier, 2000). Şekil 2.2 Leishmania parazitlerinin hayat döngüsü [1].
21 7 2.4 Leishmaniasis Dünya Sağlık Örgütü tarafından oluşturulan klinik tabloya göre, leishmaniasisin Visseral, Kutanöz, Mukokütanöz olmak üzere 3 farklı klinik formu vardır (Çizelge 2.2) (Herwaldt, 1999; Kocabas vd., 1998). Çizelge 2.2 Leishmaniasis kliniği ve etkenlerine göre sınıflandırılması (Kikuth ve Schmith, 1937) Klinik İç Organ Leishmaniasis (Visseral Leishmaniasis) Etkeni L. donovani L. infantum L. chagasi Deri Leishmaniasis (Cutanoeus Leishmaniasis) L. tropika L. major L. aethiopica L. mexicana L.braziliensisguyanensis L. b. panamensis L. peruvuiana Mukokutanöz Leishmaniasis L. b. brasiliensis Kutanöz Leishmaniasis Kutanöz leishmaniasis (KL); Şark Çıbanı, Yıl yarası, Delhi ülseri, Alleppo, deri leishmaniaisisi olarak adlandırılır. Leishmania cinsi parazitler, tatarcık tarafından kan emilen yerden derinin retikuloendotelial hücrelerine girererek amastigot formuna dönüşürler. Vektör tarafından ısırılan yerde küçük kırmızı bir papül meydana gelir, bu papül daha sonra nodül haline döner. Papüller 2 cm' den büyük çaplı, kaşıntılı lezyonlar oluştururlar ve daha sonra ülserleşerek kabuklanır. İnkübasyon periyodu 2 hafta kadar kısa (L. major) bir süre olabileceği gibi, birkaç
22 8 aydan 3 yıla kadar uzun (L. tropica, L. aethiopica) da olabilir. KL'nin Türkiye'deki başlıca etkeni L. tropica dır. Ancak son yıllarda yapılan çalışmalarda L. infantum un da KL ye neden olduğu ortaya çıkmıştır (Töre, 1996; Özçelik, 1999; Özcel, 1995; Serin vd., 2005). KL her yaşta görülebilir. Genellikle tatarcığın kan emdiği vücudun örtülmeyen kısımlarında oluşur. Örneğin yüz, ense ve kolda sıklıkla görülür. Ancak, gövdede, saçlı deride ve hatta peniste de oluştuğu bildirilmiştir. Parazit memeli konağa girdikten sonra ilk lezyonun tam olarak görülebilmesi için 2-12 haftalık bir sürenin geçmesi gerekmektedir. Daha uzun süren kuluçka süreleri de bildirilmiştir (Memişoğlu, 1994). Ayrıca dünyanın çeşitli bölgelerinde KL nin cinsiyete göre de farklılık gösterdiği bilinmektedir. Türküye de kadınların KL ile daha çok enfekte olduğu gösterilmiştir. Bunun nedeni ile ilgili çeşitli bilgiler bulunmaktadır. Ancak yaptığımız çalışmalarda bunun nedeninin kadınlarda salgılanan bazı metabolitlerin ve olduğu bu metabolitlerin Leishmania parazitlerinin üremesini stimüle ettiği gösterildi (Bagırova vd., 2009) Mukokutanöz Leishmaniasis Latin Amerika'da Espundia olarak da adlandırılan hastalığın etkeni L. Braziliensis tir. L. braziliensis ile enfekte olan kişilerde, burun, ağız, farinks ve larinkste mukozal lezyonların gelişmesi ile mukozal (mukokütanöz) leishmaniasis olarak isimlendirilen klinik tablo ortaya çıkabilir (Desjeux, 2005). Hastalığın kuluçka süresi 10 gün ile birkaç ay arasında değişebilir. Tatarcığın kan emdiği bölgede meydana gelen nodül-papül-vezikül-ülser lezyonları 10 cm'den büyük olabilir. Herhangi bir komplikasyon oluşmazsa lezyon 6 ay ile 2 yıl arasında kendiliğinden iyileşebilir. MKL lezyonları mukozal bölgelerde meydana geldiğinden, şark çıbanından ayırt edilebilir (WHO, 1990). Burundaki lezyonlar sonucu doku kayıpları oluşur. Doku harabiyeti sonucunda burun, üst dudak ve alt göz kapağında ciddi patolojik değişiklikler gelişir (Memişoğlu vd., 1994) Visseral Leishmaniasis VL etkenlerinden Leishmania donovani Hindistan ve Afrika'da, Leishmania infantum Akdeniz Bölgesi'nde ve Leishmania chagasi ise Yeni Dünya'da görülmektedir (Berman, 1997). VL yetersiz beslenme, organ nakilleri sonucu oluşan immun depresyon ve AİDS gibi immun sistemin baskılandığı durumlarda kolaylıkla gelişebilmektedir (Ak vd., 1995). Kala- Azar olarak da bilinen VL, özellikle dalak, karaciğer, kemik iliği ve lenf nodlarında bulunan mononükleer fagositik sistem hücrelerinin enfekte olması sonucu görülen bir hastalıktır. Ancak, mononükleer fagositik sistem hücrelerinin bulunduğu akciğer ve bağırsaklarda da
23 9 görülebilir (Garcia, 2001). Hastalığın kuluçka süresi genellikle 2 ila 4 ay arasındadır. Bu süre bir yıla kadar uzayabilir. Tatarcığın memeli konaktan kan emdiği yerde 2-3 mm büyüklüğünde bir nodül gelişir. Soluk pembe renkte olan bu nodülün ortası daha sonra kabuk tutmaktadır. Hastalık genellikle kendini gizleyerek sinsi başlar, immun sistemin baskılandığı durumlarda aniden ortaya çıkabilir. Başlangıçta baş ağrısı, zayıflama ve hafif ateş görülür. Daha sonra yüksek ateş ile birlikte dalak büyür. Hastalığın ilerlemesi ile ateş yükselip düşerek VL ye spesifik bir eğri çizer. Bu durum, günde iki kez yükselen aralıklı bir ateştir. Genelde C olmakla beraber bazen C'ye çıkabilir (Töre, 1996; Özbel vd., 2000) Her aralıklı ateş yükselmesinden sonra dalağın biraz daha büyüdüğü belirlenmiştir. Erişkinlerde de görülen VL genellikle çocuklarda tedavi edilmez ise ciddi enfeksiyonlara ve hatta ölümlere neden olabilir (Sundar ve Rai, 2003). Klinik olarak VL'in akut, subakut ve kronik olmak üze 3 formu olduğu belirlenmiştir (Unat vd., 1995). Subakut form; klinik olarak en sık görülen form olup akut forma göre daha belirgindir. Klinik tablosu yükselen ateş ile karaciğer ve dalağın büyümesi, kanda eritrosit, lökosit ve trombosit sayısının normalin altına düşmesi ile kendini göstermektedir. Günde 2 kez yükselip düşen ateş tipiktir, fakat hastalık sinsi olduğundan hasta genelde kendisini iyi hissedebilir. Ateş yükselişleri ile başlangıçta yumuşak olan dalağın, daha sonra sertleşip büyüdüğü gözlenmektedir. Karaciğer ilk ayın sonundan itibaren büyümeye başlar. Hastalık ilerledikçe kilo kaybı ve anemi belirgin bir hal almaktadır, fakat tüm bunlar hastanın normal hayatını etkileyecek seviyede olmamaktadır. Günümüzde yapılan çalışmalarda, hastalığın tedavi edilmesine rağmen % 1 ile % 11 arasında ölümle sonuçlandığı bildirilmiştir. Ölüm nedenleri olarak gastrointestinal sistem kanamaları, anemiye bağlı gelişen kalp yetmezliği ve karaciğer yetmezliği rapor edilmiştir. Akut form; şiddetli diş eti, burun, bağırsak kanamaları ile kemik iliği baskılanmasına bağlı oluşan kanda eritrosit, lökosit ve trombosit sayısının normalin altına düşmesi sonucu hastalık daha da ağırlaşarak ilerlemekte ve özellikle ishal şikayetlerinin eklenmesi ile hastanın genel durumu bozulmakta, hasta 2-3 ay içinde hayatını kaybetmektedir. Kronik vakalarda zayıflama, karaciğer ve dalak büyüklüğü dışında genelde hastalarda bir şikayet olmayıp, hastanın yaşamını normal olarak devam ettirdiği bildirilmiştir (Berman, 1997). 2.5 İmmunoloji Leishmania parazit enfeksiyonlarında hücresel immun yanıt ile beraber, parazitin virulans özellikleri, hastalığın gelişimi ve konağın genetik özelliklerine bağlı olarak geliştirdiği immun cevap rol almaktadır (Navin, 1988). Parazitler humoral immun cevabın oluşmasından kaçmak için, konak hücre içinde bulundukları fagolizozomlarda antijenik özellikte olan bölgelerini
24 10 değiştirerek konağı taklit etmektedirler. Enfekte makrofajlar Leishmania parazitlerine karşı nitrik oksit üretmelerine rağmen bu sistem fagolizozomlardaki parazitler tarafından inhibe edilir. fagolizozomlarda çoğalan amastigotlar konak hücreyi dağıtarak yeni fagositik hücreleri enfekte edecek olan amastigotların serbest kalmasına neden olurlar. Makrofajlar ve dentritik hücreler Leishmania antijenlerini T hücrelerine sunarlar. Sonuçta, ya hücresel ya da humoral immun yanıt oluşur. Leishmaniasis enfeksiyonuna olan direnç, Thl hücreleri ile meydana gelirken, hassasiyet Th2 hücreleri ile olmaktadır. İmmunolojik çalışmalarda parazitlerin öldürülmesinde uyarılmış makrofaj ve lenfositlerin humoral antikorlarla birlikte etkili olduğu gösterilmiştir (Schlein ve Jacobson, 1994). Ayrıca hücresel immun yanıtın, Makrofaj içindeki Leishmania parazitlerinin memeli konaktaki bağışıklığı düzenleyebileceği belirtilmiştir. İlk olarak makrofajlar Leishmania promastigotlarını fagosite ederler, daha sonra parazitler fagolizozom içinde çoğalırken ürettikleri metabolitler antijen özelliği göstererek lenfositleri uyarırlar. Bu durumda enfekte makrofajlara öldürücü etki yaparlar. Ölerek parçalanan makrofajlardan Leishmania parazitleri serbest kalır. Parazitler antikorlarla yok edilirler (Ridley, 1984). KL de hücresel immun yanıt görülmekte ve bu deri duyarlığı ile belirlenmektedir. Kendiliğinden iyileşme sonucu devamlı bağışıklık gelişir. Yapılan çalışmalarda kendiliğinden bağışıklık oluşturan hastalarda IL-2 ve IFN-γ düzeyinin arttığı, şiddetli enfeksiyonu olan olgularda ise IL-4 ve IL-10 düzeyinin arttığı gösterilmiştir (Wright ve El Amin, 1989). Deri duyarlık testi negatif olan VL de ve bazı KL olgularında, kendiliğinden iyileşme olmamaktadır. Bağışıklık türe özgü olduğundan VL etkeni L.donovani enfeksiyonu, KL etkeni L.tropica enfeksiyonuna karşı bağışıklık oluşturmaz (Manson- Bahr, 1961). Bu farklı klinik spektrum, konağın Th1, Th2, CD4 T hücrelerinin farklı cevap oluşturmasından kaynaklanmaktadır. Leishmaniasise karşı tedavinin izlenmesi açısından IFN-γ, TNF-α ve IL-6 sitokinlerinin önemli olduğu belirlenmiştir. Tedavide IFN-γ kullanılabileceği, anti-il-10 ya da inhibitor sitokinleri baskılayıp aynı zamanda da IFN-γ sentezleten IL-12 gibi sitokinlerin kullanılabileceği düşünülmektedir. Aşı çalışmalarında ise, IL-12 gibi immun sistemi güçlendirecek, sitokinlerin sentezini uyaracak antijenik özellikteki parazit proteinlerinin kullanılması ile etkili sonuçlar alınabileceği düşünülmektedir (Ak vd., 1995; Berman, 1997). Akut VL'de ilk 4 hafta IL-2 ve IFN-γ sentezlenmediği, ancak tedaviden sonra salgılandığı gösterilmiştir. Subakut VL de ise IFN-γ sentezinin çok az olduğu görülmüştür. Akut VL'li hastalarda IL-2 reseptörü, kanda serbest olarak bulunan IL-2 ile birleşerek hedef hücreye
25 11 ulaşmasını engellemektedir. (Wright ve El Amin, 1989). Akut VL'li hastaların lenf nodüllerinde çok yüksek oranda IL-10 mrna sentezi olduğu ancak tedavi edilen hastalarda sentezin durduğu belirlenmiştir. L.mexicana nın neden olduğu KL hastalarında, IL-10 ve TGF-α da dördüncü aydan itibaren belirgin bir artış görülür ve makrofajlar üzerine inhibitör etki yaparak enfeksiyonun kronikleşmesine neden olur (Ak vd., 1995). Bazı Leishmania türlerinde makrofaj fonksiyonlarını inaktive eden TGF-α ve IL-10 gibi sitokinler, hastalığın patogenezinde önemli rol oynamaktadır. Parazitin antijenik yapısı üzerinde yapılan çalışmalarda, genelde somatik yapıda olan 30 kadar antijenin olduğu belirlenmiştir. Sonuç olarak parazitin antijenik yapısı ve bu antijenlere karşı oluşan hücresel ve humoral immun cevaplara yönelik çalışmalar ile leishmaniasise karşı aşı geliştirme çalışmaları hala devam etmektedir Leishmania Parazitlerinin Konak Hücre İçindeki Yaşamı Metasiklik promastigotların makrofajlara fagosite edilmesi ve hayatta kalması, parazitin sitotoksik serum komponentlerinden kaçmasına, makrofaj içersinde hayatta kalmasına ve T hücre immun cevabı oluşturmamasına bağlıdır (Chang vd., 1985). Tatarcıkların kan emmesi sırasında promastigotlar, mononukleer fagositik hücreler tarafından fagosite edilir. Hücre içine giren promastigotlar, amastigot forma dönüşürler. Yapılan in vitro çalışmalar promastigot yüzey membranında bulunan üç molekülün enfektiflik açısından önemli olduğunu göstermiştir. Bunlardan lipofosfoglikan (LPG), glikokaliksi oluşturur ve glikokaliks üzerinde bulunan C 3 b, promastigotun konağın kompleman sistemi tarafından imha edilmesini önlediği gibi aynı zamanda konak hücreye girmesine ve parazitin makrofaj içinde hayatta kalmasını sağlar. Promastigotlar komplemanı klasik yoldan aktive eder ve konak kompleman reseptörlerini konak hücreye girmek için kullanır. Yani promastigotlar makrofajlar tarafından reseptör temelli fagositozla içeri alınır. fagositozla hücre içine alınan promastigotların bulunduğu fagolizozom oldukça asidik (ph: 4,5-5.0) ve proteazlara sahip olmasına rağmen parazitler burada yaşamını sürdürebilir. Lipofosfoglikan Leishmania türleri arasında farklılık gösterir ve promastigotlar tarafından kültür ortamında da salgılandığı için Leishmania tür tayininin yapılmasında kullanılır. Diğer iki molekül ise major glikoprotein gp63 ve asit fosfatazdır (Awasthi vd., 2004; Chang vd., 2002; De Almeida vd., 2003). LPG ve gp63, adezyon moleküllerini (Hücre içi Adezyon Molekülü (ICAM), Vasküler Adezyon Molekülü (VCAM) ve Makrofaj Kemotaktik Protein (MCP-1)) inhibe ederek makrofaj göçünü engeller, oksijen radikalleri ile nitrik oksit baskılanması için protein kinaz C
26 12 sinyal yolunu inhibe ederek, parazitin hidrolitik enzimlerle hücre içi parçalanmasını engeller. Ayrıca fagolizozom oluşumunu geciktirerek, promastigotların amastigot formuna dönüşmesini ve amastigotun diğer hücreleri enfekte etmesi için zaman kazanmasını sağlar (Alexander vd., 1999). 2.6 Leishmania Parazitlerinin Kültürünün Yapılması Leishmania parazitlerinin in vitro kültürünün yapılması parazitlerin biyokimya, immünoloji ve hücre biyolojisi üzerine çalışma yapılmasına olanak sağlar. Memeli konaklarda hücre içinde yaşayan amastigotların kültürü, makrofaj benzeri hücreler içinde yapılmasına nispeten zor ve zahmetli olması nedeniyle, promastigot kültürü tercih edilmektedir. Yine de makrofaj hücrelerinde bulunan amastigotların pasajları yerine, aksenik amastigot kültürleri de başarıyla yapılabilmektedir (Chang ve Bray, 1985a; Chang ve Bray, 1985b; Gholamhosseinian ve Vassef, 1988). Promastigot ve amastigot formdaki parazitlerin büyüme koşulları arasında, sıcaklık, ph, besin kaynağı ve serum komponentleri açısından önemli farklılıklar bulunur. Sıcaklık ve ph ın, promastigotların amastigotlara dönüşümünü tetiklemede önemli rolü vardır. Bu şekilde ortam özelliklerini değiştirerek promastigotlar aksenik amastigotlara dönüştürülebilir (Somana vd., 2002). Leishmaniasisli hayvanlardan alınan biyopsi örnekleri, uygun besiyerine ekilerek Leishmania promastigot kültürü yapılabilmektedir. Kültüre ekilen biyopsi örneklerindeki Leishmania lar önce hareketli promastigotlara dönüşür daha sonra çoğalmaya başlar (Chang ve Bray, 1985a; Chang ve Bray, 1985b) Kullanılan Besiyerleri Leishmania nın in vitro kültürünün yapılmasında birçok besiyeri kullanılmaktadır. Bu besiyerlerinin her birinin farklı avantaj ve dezavantajları vardır. Bugün, Leishmania'nın ürediği besiyerlerinin tümü, diğer hücrelerin üredikleri besiyerlerinin denenmesi ile bulunmuştur. (Chang ve Bray, 1985a; Chang ve Bray, 1985b; Gholamhosseinian ve Vassef, 1988) yılında, McNeal ve Novy tarafından Trypanosoma kültürünü elde etmek için hazırlanan bifazik besiyeri, Nicole tarafından modifiye edilerek Leishmania promastigotlarını üretmek için denenmiş ve tüm dünyada Novy-MacNeal ve Nicole (NNN) ismiyle bilinen besiyeri geliştirilmiştir (Chang ve Bray, 1985a; Saran vd., 1986; Gholamhosseinian ve Vassef, 1988).
27 13 Günümüzde Leishmania kültürü için çeşitli katı, yarı katı ve sıvı besiyerleri bulunmaktadır. Katı besiyerlerinden olan NNN besiyeri; agar, tuz ve defibrine edilmiş tavşan kanından oluşmaktadır. Besiyerine ayrıca %20 fetal dana serumu (FDS) ilave edilerek 5 14 günde promastigotların üremesi sağlanmıştır (Charles ve McMahan, 1984; Bard, 1985). Fetal dana serumu kullanılan besiyerlerinde, Leishmania %25 oranında daha fazla üremektedir. Besiyerindeki besinin hızla tükenmesi, hücre ölümüne neden olacağından, Leishmania kültürünün sık sık rutin pasajlarının yapılması gerekmektedir (Sadigursky ve Brodskyn, 1986). Schneider'in besiyeri ise sıvı bir besiyeri olup birçok amino asit içermektedir. Bu besiyeri genelde immunolojik ve biyokimyasal çalışmalar için kullanılmaktadır. Ayrıca belli oranlarda FDS eklenmesi ile promastigot kültüründe kullanılması uygundur (Charles ve McMahan, 1984). Schneider'in besiyeri, Leishmania'nın üremesini hızlandırmakta böylelikle kısa sürede yüksek parazit miktarlarına ulaşılabilmektedir (Chang ve Bray, 1985b). Daha çok tanı amaçlı kullanılan beyin-kalp infüzyonu kan eriyiği, NNN besiyerinden agarın çıkartılması ile hazırlanmakta ve kısa sürede bol miktarda Leishmania üretilmesini sağlamaktadır (Gholamhosseinian ve Vassef, 1988). Ayrıca tampon olarak HEPES kullanılan ve genelde %10 20 FDS eklenmesi ile uygun sonuç alınabilen Medium 199 ile Leishmania pasajı 2 haftaya kadar dayanabilmektedir (Chang ve Bray, 1985b). Bunun yanı sıra RPMI 1640 gibi besiyerlerine %10 FDS eklenmesi ile rutin promastigot kültürü başarılı bir şekilde yapılmaktadır. Biyokimyasal çalışmalarda, sıvı besiyeri önerilirken, izolasyon ve pasajlarda modifiye NNN besiyerleri önerilmektedir (Chang ve Bray, 1985b; Rao vd., 1984). Sıvı besiyerleri ile kültür yapılmasının birçok avantajı vardır. En önemlisi kapalı bir sıvı örnek içersinde birçok hareketli promastigot bir arada toplanıp çalışılabilmektedir. Hastalık tanısı konulmasında, sıvı besiyerleri özellikle parazit sayısının yüksek olduğu örneklerde oldukça avantajlıdır. Ayrıca Allahverdiyev vd. (2004) Dünya da ilk kez olarak daha hassas, daha spesifik, daha duyarlı, daha hızlı ve daha ekonomik olan Yeni Mikrokültür Yöntemi geliştirilmiştir. Klsasik yöntemden farklı olarak hastalığın tanısı 1-6 ay yerine en fazla 2 gün içinde konulabilmektedir (Allahverdiyev vd., 2005) Bu yöntyem Dünya nın çeşitli bölgelerinde uygulanmaktadır (Boggild vd., 2008). Bu yöntem Mikrokültür yöntemi olarak bilinen bu yöntemde sıvı besiyeri ortamında parazitler konsantre edilip, mikroaerofilik bir ortam sağlanarak, kısa sürede amastigotların promastigotlara dönüştüğü ve bu promastigotların hızlı bir şekilde çoğaldığı gösterilmiştir.
28 14 Yapılacak çalışmaya uygun besiyeri seçilip, besiyerlerinin ph'ının: 6.8 ila 8'e arasında ayarlanması gerekir. İnkübasyon derecesi ise genellikle C arasında olmaktadır. Ancak promastigotların bazı durumlarda 38 C'ye de uyum sağlayabildikleri görülmüştür (Chang ve Bray, 1985b). Makrofaj içinde yapılan amastigot kültürü için CO 2 etüvü gerekirken, promastigot kültüründe böyle bir gereksinim yoktur, sadece sıcaklığın ayarlanması yeterlidir. Promastigotların kültüründe farklı plastik ve cam tüpler kullanılabilmektedir. Kullanılan tüpler veya flasklar, vidalı kapaklı olup hava geçirmeyecek şekilde kapalı tutulur. Rutin pasajlar, eski kültürden alınan promastigotun, yeni besiyeri içine konulmasıyla yapılır. Bakteri kontaminasyonlarını önlemek için besiyeri içine Penicillin, Streptomycin veya Gentamycine gibi antibiyotikler katılabilmektedir (Alfred ve Steve, 1986). Rutinde kullanılan bu antibiyotikler, promastigotların üremesini etkilememektedir. Ayrıca, Leishmania kültürlerindeki mantar kontaminasyonlarını önlemek amacıyla Mycostatin ve Amfotericine B gibi antifungal bileşikler besiyerine eklenebilmektedir (Charles ve McMahan, 1984). Leishmania kültüründe kontaminasyon oldukça önemli bir problemdir. Bu sebepten kültürün steril ortamda yapılması zorunludur (Charles ve McMahan, 1984; Jackson vd., 1989). İn vitro ortamda çoğalan tüm Leishmania promastigotlarının ortak özelliği; diğer organizmalarda olduğu gibi logaritmik üreme ve sabit faz dönemlerinin olmasıdır. Genellikle ilk inokülasyonda hücre/ml olan parazitler, 3-4 günlük inkübasyonun aradından hücre/ml'ye kadar çıkabilmektedir (Sadigursky ve Brodskyn, 1986). Rutinde, örnek sayısı çok olduğu zaman hücrelerin sayımında Coulter sayacı kullanılabilir. Ayrıca, Leishmania'nın mikroskobik incelenmesi invert mikroskopla yapılabilmektedir (Bray, 1985). Kültür süresince, kültürdeki promastigotlar sıkça pleomorfizm gösterirler. Birçok türde promastigotların eni µm, boyu ise 1-20 µm arasında değişebilir (Gholamhosseinian ve Vassef, 1988). Yuvarlak, oval, eliptik veya silindir şekilde olabilirler. Bazen de uç kısımları sivrilebilir veya yuvarlaklaşabilir. İyi hazırlanmış bir kültürde, kendiliğinden hareket edebilen ve bölünen hücrelerin olması gereklidir. Çok sayıda hareketsiz ve normalden büyük veya aşırı küçük atipik hücrelerin varlığı, kültür ortamının yetersizliğinin göstergesidir (Chang ve Bray, 1985b). Primer izolasyonda önemli olan, en uygun besiyerine karar vermektir çünkü bazı Leishmania türleri ilk izolasyonun yapıldığı besiyerinin değiştirilmesine adapte olamazlar. Eğer, besiyerleri arasında karşılaştırma yapılmak isteniyorsa, ilk besiyerine uyum sağlandıktan sonra alınan örneğin, birden fazla farklı besiyerlerine ekilmesi gerekir. Leishmania'nın primer
29 15 olarak izolasyonu sırasında %10 FDS eklenmiş RPM besiyeri kullanılabilmektedir (Chang ve Bray, 1985b; Alfred ve Steve, 1986; Gholamhosseinian ve Vassef, 1988). Promastigotların üretilmesi, amastigotlara oranla biraz daha kolaydır. Kamçısız amastigotlar, ancak memeli makrofaj hücrelerinde üremektedir. Amastigotların ısı tercihi de promastigotlardan farklı olup 32 C dir (Alfred ve Steve, 1986). Leishmania amastigotlarının üremesi konusunda başarılı deneysel çalışmalar yapılmış, memelilerden hazırlanan doku kültürlerinde hücre içinde yaşayan amastigotların araştırılması kolaylaşmıştır (Chang ve Bray, 1985a; Chang ve Bray, 1985b; Gholamhosseinian ve Vassef, 1988). Doku kültüründeki teknik ilerlemeler, amastigotların in vitro kültürünü de olumlu yönde etkilemiştir. Sonuçta memelilerde hücre içi üretilen amastigotların ardından bu yöntem diğer Trypanosoma'lara da uygulanmış ve başarı sağlanmıştır (Ashford ve Betini, 1987). Bu konuda yapılan ilk araştırmalar, amastigotların embriyoda üretilmesi ile ilgili olup, karaciğerde az miktarda amastigotların üremesiyle başlamıştır. Halbrook ve Palcuk 1975'de Leishmania promastigotlarını tavuk embriyosunda üretmeyi denemişler ve embriyonun karaciğerinde kısa bir sürede üreyebilen amastigotları elde edebilmişlerdir (Chang ve Bray, 1985b). Bugün, çeşitli hayvanlardan alınarak hazırlanan primer makrofaj kültürleri, araştırmalarda amastigotların standart kültür sistemini oluşturmaktadır (Alfred, 1983). Bu sistemler sayesinde, laboratuar hayvanlarına bağlı kalmaksızın Leishmania'ların yaşam döngüleri ve amastigotların biyokimyasal analizleri araştırılabilmektedir. Yapılan bu kültür yöntemiyle, amastigotların devamlılığı da sağlanmıştır. Leishmania türleri; hazır besiyerlerinde üretildikleri gibi, deney hayvanlarına inoküle edilerek de üretilebilirler. Deney hayvanları kullanılarak; Leishmania ların vektörlerdeki yapısı, biyolojisi ve patolojisi deneysel olarak araştırılabilmektedir (Chang ve Bray, 1985b; Grimaldi vd., 1989). Günümüzde de enfekte edilmiş deney hayvanları kullanılarak aksenik amastigotlan elde etme çalışmaları devam etmektedir (Scott vd., 1987). Leishmania türlerinin devamlılığı, yapılan promastigot ve amastigot kültürleri ile hayvanlara inokülasyonla ya da diğer memeli hücreleri veya protozoanlarda olduğu gibi sıvı azotta dondurup uzun süre saklayarak sağlanabilir (Chang ve Bray, 1985b; Grimaldi vd., 1989).
30 16 3. Mononükleer Fagositik Sistem Hücrelerinin Gelişimi Mononükleer fagositik sistem, fonksiyonu fagositoz olan hücrelerden oluşmuş olup immun sistemin ikinci büyük hücre grubudur. Bu sistemin bütün hücreleri kemik iliğinden gelişir (Şekil 3.1) Kemik iliğinde bulunan öncü hücrelerden, multipotent miyeloid hücreler oluşur. Bu hücrelerden ise monosit colony forming unit (CFU-M) hücreleri meydana gelir. CFU-M hücreleri monoblastlara dönüşür. Monoblastlarda bölünerek promonositleri oluşururlar. Promonositler monositlerden daha küçük hücreler olup µm'lik çapa sahiptirler. Kemik iliğinde bulunan promonositlerin yaklaşık yarısı hızlı bir şekilde bölünüp, küçülerek bölünebilme özelliğini kaybeden monositlere farklılaşırlar. Geriye kalanlar promonositler ise gerektiği zaman monositlere dönüşürler. Kök hücreden monosite farklılaşma yaklaşık olarak 55 saat olup bu monositler kemik iliğinde 1-2 gün kaldıktan sonra kana geçerler (Janeway vd., 1999; Junqueira vd., 2003). Monositlerin çapları yaklaşık µm olup büyük bir çekirdeğe sahiptirler. Çekirdekleri merkezden içe doğru çöküntü oluşturarak böbreğe benzemektedir. Bir veya iki çekirdeğe sahip olabilirler. Monosit sitoplazmasının iyi gelişmiş Golgi kompleksi, ribozom ve poliribozomlar, çok sayıda küçük mitokondri içerdiği belirlenmiştir. Az sayıda granüllü endoplazmik retikulum içerirler. Hücrenin periferinde mikrotübül ve mikrofilamanlar, pinositik veziküller bulunur. Hücre yüzeyinde mikrovilluslar vardır. Monositler dolaşımda birkaç gün kalıp bağ dokuya geçerek makrofajları oluşturmaktadır (Junqueira vd., 2003; Eşrefoğlu, 2004). Monosit ve endotel hücre membranlarındaki hücre adhezyon molekülleri monositlerin endotele tutunarak yavaşlamasını, durmasını ve endotel hücreleri arasından dokulara geçişini sağlar. Bu göç sırasında ilk adım kan damarlarında dolaşan monositin, endotel hücresinin yüzeyine tutunmasıdır. Daha sonra damar yüzeyinden yavaşça yuvarlanarak sürüklenir ve daha güçlü bir şekilde tutunarak durur. Bu olayda selektinler, β-1 integrin ve endotel yüzeyindeki VCAM-1 gibi adhezyon molekülleri ilk tutunmada görev alırken, lökosit β-2 integrin ile ICAM-1 ve 2 ise monositin durmasını sağlayan adhezyon molekülleridir. Monositler daha sonra endotel hücreleri arasından, hem monosit hem endotel yüzeyinde bulunan PECAM-1 molekülleri aracılığı ile dokuya geçerler. Endotel bazal membranını geçen monositler makrofajlara dönüşecekleri bölgelere göç ederler (Maslin vd., 2005).
![17 Şekil3.1 Makrofaj aktivasyonu [2] 3.1 J774 Hücre Hattı J774 fare makrofaj hücre hattı dişi bir BALB/c farede ortaya çıkan tümörden elde edilmiştir.](/docs-images/67/57510468/images/31-0.jpg "Hücre hattının çoğalması dekstran sülfat, bakteriyel lipopolisakkaritler ile inhibe olur.")
31 17 Şekil3.1 Makrofaj aktivasyonu [2] 3.1 J774 Hücre Hattı J774 fare makrofaj hücre hattı dişi bir BALB/c farede ortaya çıkan tümörden elde edilmiştir. Hücre hattının çoğalması dekstran sülfat, bakteriyel lipopolisakkaritler ile inhibe olur. J774 hücre hattı monosit-makrofajların fizyolojisini ve makrofaj koloni sitimüle edici faktörün (M- CSF) fonksiyonunu anlamak amacıyla birçok biyokimyasal çalışmada kullanılmıştır. Bu hücre hattı yüksek miktarda lizozim sentezler ve çok az sitoliz görülür. Fakat genellikle antikor bağımlı fagositoz meydana gelir. J774 hücreleri, IFN-γ çok düşük konsantrasyonlarında bakteriyel lipopolisakkarit (10 ng/ml) ile aktive olduğu zaman nitrik oksit sentaz enzimi yüksek seviyede aktive olduğu için büyük miktarlarda nitrik oksit üretir. IFN-γ, J774 hücrelerinde IL-12 nin sentezlenmesini indükler. J774 hücre hattı hücre kültür ortamlarına sürekli olarak IL-1 salgılar [3]. 3.2 Makrofajların Aktivasyonu ve Fonksiyonları Monositler dokulara geçip makrofaja dönüştükten sonra yaklaşık 2-4 ay arasında yaşarlar. Bu süre içinde bazı makrofajlar dokuya tutunarak hareketsiz kalırlar. Bunlara sabit makrofaj denir. Genellikle matriksteki kollajen fibrillere tutunurlar. Geriye kalan makrofajlar ise ameboid hareketle sürekli yer değiştirirler. Bütün doku makrofajları yüzeylerinde bulunan reseptörlerle ve pinositoz yaparak çevrelerini denetlerler. Eğer çevreden gelen herhangi bir sinyalle veya antijenle karşılaşırlarsa makrofaj aktivasyonu gerçekleşir. Bu süreçte
32 18 makrofajların büyüklükleri, metabolik hızları, hareketlilikleri ve fagositik aktiviteleri hızla artar. Hücre Büyüklüğünün artması sitoplazmik hacmin artmasıyla olur. Aktivasyonla pek çok yeni madde sentezlenir. Bunlar içinde en önemli olanlarından biri indüklenebilir nitrik oksit sentazdır (İNOS). Bu enzim ile L-arginin den son olarak oluşan nitrik oksit (NO) çok düşük konsantrasyonlarda bile mikroorganizmalar ve tümör hücreleri için sitotoksik etkiye sahiptir (Abbas vd., 1994). Makrofajların belli başlı önemli fonksiyonları şunlardır: Fagositoz Makrofajlar tümör hücreleri, parazit, bakteri, makromolekül, antijen gibi yabancı maddelerle, hasarlı, ölü hücre ve artıklarını fagosite ederler. Makrofajların hücre membranında farklı antijenik maddeler için spesifik reseptörler bulunur. Makrofajların yabancı partikülleri ve hasarlı dokuları tanımasında fosfolipid ve şeker reseptörlerinin önemli rolü vardır. Bunlar içinde mannoz reseptörü, membran CD 14 reseptörü, toll-like reseptör (TLR) ailesi sayılabilir (Abbas vd., 1994). Makrofajlar, antikorları ise yüzeylerinde bulunan Fc reseptörleri ile tanırlar. Fagosite edilecek madde yüzey reseptörlerine bağlanır ve bu bağlanma ile sindirilecek maddenin hücre içine alınmasıyla sitoplazmada fagozom oluşur. Fagozomlar hücre içerisinde bulunan lizozomlarla birleşerek fagolizozomu (sekonder lizozom) oluşturur. Lizozomal enzimler sindirilecek materyalin komponentlerinin katabolik reaksiyonlarla yıkılıp, sitoplazmaya verilmesini sağlar. Ayrıca makrofajlarda moleküler oksijen indirgenerek üretilen reaktif oksijen radikalleri ile hücre içi enfeksiyon ajanlarını öldürürler. Fagolizozom içinde oluşan komponentler ve artık maddeler ekzositoz yolu ile hücreden atılır veya sitoplazma içerisinde artık olarak birikir (Parslow vd., 2001). Sindirilecek materyalin fazla sayıda, büyük veya sindirilmeye dirençli olmasına bağlı olarak bölgeye çok sayıda makrofaj birikir. Ayrıca bu bölgelerde lenfositler, fibroblastlar ve diğer hücreler de birikir. Bu şekilde oluşan yapılara granüloma denir. Granülomadaki makrofajlara epiteloid hücreler denir. Bunun nedeni makrofajların epitel hücrelerine benzer olarak yan yana gelerek, hücre-hücre bağlantıları oluşturacak şekilde birbirlerine uzantılar göndermeleridir. Böylece granüloma dışına madde geçişini engellenmiş olur (Parslow vd., 2001). Dokulara giren yabancı bir cismin fagositozla alınması tek bir hücre için çok zor olduğu zaman birçok makrofaj bir araya gelip birleşerek çok çekirdekli dev bir hücre yapısı oluştururlar. Buna yabancı-cisim dev hücresi denir. Böylece büyük zararlı-yabancı cisimler yakalanıp, sindirilebilir (Young vd., 2001).
33 Salgılama Makrofajlar biyolojik olarak aktif olan çok fazla madde salgıladığı ve bu salgılanan maddelerin bugüne kadar 100'den fazla çeşidi belirlenmiştir. Bunlardan lizozim, hidrojen peroksit gibi maddelerin antimikrobiyal aktiviteleri varken, elastaz ve kollajenazlar ekstrasellüler matriks döngüsünde görev alırlar. Makrofajlar ayrıca sitokin adı verilen sinyal molekülleri salgılayarak immun sistemi regüle ederler. Sitokinler, immun sistem hücreleri arasında iletişimi sağlayan küçük proteinlerdir. En önemlileri arasında TNF α, interlökin-1β, interferon α, β, interlökin 6, 10, 12, fibroblast büyüme faktörü (FGF), prostoglandinler, kemokinler, nitrik oksit vb. bulunmaktadır (Abbas vd., 1994) Antijen Sunma Makrofajlar antijenlerle birleşen ilk hücreler olup, antijen sunma özellikleri vardır. Yabancı maddelerle birleşerek onların lenfositler tarafindan taninmalarını ve lenfosit cevabı oluşmasını sağlarlar. Yüzeylerinde major histokompabilite kompleks II (MHC II) olarak adlandırılan spesifik proteinler içerirler. Bu moleküller antijene spesifik T-helper (CD4 + ) lenfositler tarafından tanınır. Makrofajlar, yabancı bir hücreyi veya antijeni fagosite ettikten sonra elde ettikleri antijenler MHC II moleküllerinin yüzeyine tutunur. Eğer CD4 + T lenfosit bu antijeni tanirsa aktive olur ve immun reaksiyon başlar (Parslow vd., 2001). 3.3 Makrofajların Özellikleri Makrofajların yapısal özellikleri aktif olma durumlarına ve dokudaki yerleşimlerine göre oldukça farklılık gösterir. Düzensiz şekilli ve çeşitli büyüklüklerde olabilen makrofajlar genelde µm çapındadır (Parslow vd., 2001). Makrofajlar Işık mikroskobunda, ancak fagosite edebilecekleri boyalar verildiğinde görünür olurlar. Çekirdekleri oval veya böbrek şeklinde görünürken çekirdekçik genellikle görülmez. Fagositik özelliğe sahip oldukları için sitoplazmalarındaki en belirgin organel lizozomlardır. Bunun dışında Golgi kompleksi, granüllü ve granülsüz endoplazmik retikulum, mitokondri, sekretuar veziküller, endositik veziküller ve fagolizozomlar içerirler. Makrofajların yüzeylerindeki çıkıntılar, içerdikleri bol lizozom, endositik veziküller, fagolizozomlar ve reziduel cisimler fagositik aktivitelerinden kaynaklanır (Young vd., 2001).
34 20 Makrofajlar aktifleşirken protein sentezi artar ve hücreler normalden daha büyük olur. Lizozomların, mikrotübüllerin ve mikrofilamanların sayılarıdaki artışın yanı sıra Golgi kompleksinde de bir artış görülür. Granüllü endoplazmik retikulum ve Golgi kompleksi fagositoz için gerekli proteinlerin sentezini ve salgı ürünlerinin sentezini sağlar. Lizozomlar fagositozun yapıldığı organeller olduğundan, sitoplazma içerisinde bol miktarda bulunurlar. Golgi kompleksi tarafından oluşturululan lizozomlar hücreye giren yabancı maddeleri sindirir, hücre metabolizma artıklarını parçalarlar. Herhangi bir sindirim işleminde hiç görev almamış olan lizozomlara primer lizozom denir. Primer lizozomlar daha küçük olup ince granüler ve homojen içeriğe sahiptir. Sindirimde görev aldıktan sonra sekonder lizozom adı alırlar. Sekonder lizozomlar büyük olup içyapıları değişikliğe uğrayarak heterojen özellik göstermeye başlar (Junqueira vd., 2003; Eşrefoğlu, 2004). 3.4 Nitrik oksit (NO) Nitrik oksit (NO) yarı ömrü kısa ve gaz yapısında bir moleküldür (Grisham, 1997). Nitrik oksit, üzerinde yük taşımaması, çiftleşmemiş elektron bulundurması ve lipofilik özellikte olması hücreden hücreye hiçbir bariyerle karşılaşmadan kolaylıkla geçmesini sağlar. Oksijensiz ortamda oldukça stabildir. Düşük konsantrasyonlardayken, ortamda oksijen olsa bile oldukça kararlı olan NO, bilinen en düşük molekül ağırlıklı, biyolojik olarak aktif memeli hücresi salgısıdır. Diğer serbest radikaller hücreler için zararlı iken NO düşük konsantrasyonlarda bile önemli fizyolojik islevlerde rol alır. NO bu özellikleri ile ideal bir fizyolojik haberci molekülü özelliğine sahiptir (Lowenstein vd. 1994). INF-γ veya bakteri lipopolisakkaritleri tarafından aktive edilen makrofajlarda, nitrik oksit sentaz (NOS) tarafından L-arjininden sentezlenen NO, intrasellüler parazitlere karsı çok önemli bir savunma mekanizmasında rol oynar (Chiueh 1999; Ghafourifar ve Cadenas 2005). Nitrik oksit sentazın, nöronal NOS (nnos), endoteliyal NOS (enos) ve indüklenebilir NOS (inos) olmak üzere üç farklı izoenzimi vardır. İlk iki formu konstütif olup, kalsiyuma bağlı olarak endoteliyal NO in üretiminden sorumlu iken inos kalsiyumdan bağımsızdır ve büyük oranda makrofaj kökenli NO sentezinden sorumludur (Ahren vd., 1999, Lopez-Urritia vd., 2000). L-Arginin'e etki eden iki enzim vardır. Bunlardan birincisi Nitrik Oksit Sentaz-ll (NOS-II) olup makrofajlar tarafından salgılanarak nitrik oksit (NO) üretilmesini sağlar. Diğer enzim ise arginaz olup görevi L-argininden L-ornitin ve üre oluşumunu sağlamaktır. NOS-II enziminin son ürünü olan NO, mikromolar konsantrasyonlarda bile mikrobik organizmalar ve tümör hücreleri için sitotoksik etkiye
35 21 sahiptir. Arginaz enziminin son ürünü olan L-Ornitin ise özellikle parazit gelişimi için faydalı olduğu bildirilmiştir. Makrofaj kaynaklı NO Leishmania parazitleri üzerinde sitotoksik etki yapar. Yüksek NO, bakterilerde DNA sentezinin hız kısıtlayıcı enzimi olan ribonükleotid redüktazı bloke ederek hücre DNA sının deaminasyonu ile sitotoksik etki yapar (Lin ve Chadee 1992, Taylor- Robinson ve ark 1996). inos ekspresyonu normal fizyolojik şartlar altında gerçekleşmezken herhangi bir yabancı madde ya da hücrenin vücuda girmesi durumunda sitokinler veya endotoksinler tarafından indüklenir ve böylece uzun süre yüksek konsantrasyonlarda NO üretilebilir. inos basta makrofajlar olmak üzere polimorfonükleer lökositler, nötrofiller, hepatositler, düz kas hücreleri ve kondrositler gibi birçok hücrede uyarılabilir. inos tarafından üretilen NO makrofajlarda antileishmanial etkiye sahiptir ve bu nedenle nonspesifik konak savunma sisteminin önemli bir parçasıdır (Vincent vd., 2004, Pratic`o 2005). 4. Polimerler 4.1 Polimerlerin Genel Özellikleri ve Sınıflandırılmaları Polimerler monomer adlı basit yapıların birleşmesiyle oluşan uzun zincirli moleküllerdir. İki veya daha fazla monomerin birleşiminden yapılabildikleri gibi tek bir monomerden de yapılabilirler (Chandrasekhar, 2005). Polimerlerin, farklı kriterleri göz önünde bulundurularak, farklı sınıflandırmaları yapılmaktadır. Örneğin molekül ağırlıklarına göre (oligomer, makromolekül), doğal veya sentetik olmalarına göre, ısıya karşı gösterdikleri davranışlara göre (termoplastikler, termosetler), zincirin kimyasal ve fiziksel yapısına göre (düz, dallanmış, çapraz bağlı, kristal, amorf; homopolimer, kopolimer), sentezlenme şekillerine göre sınıflandırılabilirler. Polimerleşme reaksiyonları esnasında pek çok monomer, diğer monomerlerle ya da ortamda daha önce tepkime vermiş ve belli bir moleküler ağırlığa ulaşmış molekül zinciri ile tepkime verebilir. Eğer polimer zinciri yeterince büyümemişse oligomer olarak adlandırılır. Oluşan zincirlerin büyüklükleri, türlerin moleküler yapılarından, tepkime verme yollarına ve sentez şekillerine kadar pek çok faktöre bağlıdır. Uzun zincirler halinde bulunan düz ya da dallanmış zincirlerden oluşan polimer sistemlerine termoplastik adı da verilmektedir. Yüksek oranda çapraz bağ içeren sistemler ise termosetting olarak tanımlanır. Termoplastik bir malzeme, sıcaklık artışı ile eriyerek şeklini değiştirebilir
36 22 ve böylece kalıplara dökülebilmesi mümkün olur. Oysa yüksek oranda çapraz bağ içeren bir yapının sıcaklık artışı ile böyle bir değişim geçirmesi, zincirlerin hareketli olmayışı nedeniyle, zordur ve sıcaklığın artışı bu tür malzemenin erimesine değil, parçalanmasına neden olur (Baysal, 1994). Polimerlerin çoğu etilen, stiren gibi basit monomerlerden hazırlanır. Poli(etilen terethalate) veya naylon 6,6 gibi polimerler iki monomerin reaksiyonundan hazırlanırken, iki monomerden fazlasının reaksiyonunu içeren polimerler de vardır. Buna örnek olarak akrilonitril, stiren ve butadienin polimerizasyonuyla oluşan poli(akrilonitril-butadien-stiren) kopolimeri verilebilir (Chandrasekhar, 2005). Sentetik polimerlerin yanı sıra doğal olarak oluşan birçok polimer vardır. En iyi bilinen doğal polimerler; proteinler, deoksiribonükleik asit (DNA), hemoglobin, insülin, nişasta gibi yapılardır (Chandrasekhar, 2005). Polimerler iyon içeriklerine göre homopolielektrolitler ve poliamfolitler olmak üzere iki grup oluşturmaktadırlar. 4.2 Polielektrolitler Polielektrolitler, kovalent olarak bağlı anyonik veya katyonik grupları ve bu gruplara bağlı iyonları bulunan polimer sistemleridir. Polielektrolitler, üzerinde bulunan yüklü grupların tümünün eksi yüklü olması durumunda polianyon adını alırken, tümü artı yüklü ise polikatyon adını alır. Poliamfolitler yapılarında kovalent bağlı olarak anyonik ve katyonik grupların her ikisini de içeren makromoleküllerdir (Şekil 4.1).
37 23 Şekil 4.1 Polielektrolitlerin sınıflandırılması a.polikatyon b.polianyon c.poliamfolit (Koetz, 2007) Polifosfatlar ve polisilikatlar inorganik polielektrolitler olarak sayılabilir. Sentetik polielektrolitler genelde bir katılma veya kondenzasyon polimerleşme reaksiyonlarıyla elde edilebilir. Sentetik yapıda polielektrolitlerin dışında, polisakkaritler, proteinler ve nükleik asitler gibi doğal polielektrolitler de mevcuttur (Hileman vd., 1998). Bunların yanı sıra karboksimetilselüloz gibi yarı sentetik polielektrolitler, doğal polimerlerin özelliklerinin iyileştirilmesi için modifiye edilmesiyle elde edilir (Trabelsi ve Langevin 2007). Polielektrolite bağlı elektrolit gruplar sulu çözeltilerde dissosiye olup polimeri yüklü hale getirirler. Bu durum polimerin konformasyonunu değiştirir. Herhangi bir polimerin konformasyonu, polimerin yapısı ve çözeltiyle ilişkisi gibi çeşitli etkenlere bağlıdır. Çözelti içindeki yüksüz, düz bir polimer zinciri rasgele yumak konformasyonu gösterirken, polielektrolitler üzerindeki yüklerin birbirini itmesi sonucu uzanmış, düz bir konformasyon oluşturmaya eğilim gösterir (Coulomb etkisi). Bu konformasyon, polielektrolit çözeltilerinin viskozitesinin yükselmesine yol açar. Polielektrolit çözeltilerinin özellikleri, sulu ortamın iyonik şiddetine bağlıdır. İyonik şiddetin artmasıyla polielektrolitteki iyonların elektrik yükleri arasındaki itme kuvvetinin engellenmesi viskozitenin değişmesine neden olur. Polielektrolitler, çözeltideki iyonik şiddetin daha fazla artması ile yüksüz polimerlere benzer davranış göstermeye başlarlar (Bromberg, 2001).
38 24 Zayıf polielektrolitlerin özellikleri iyonik şiddetin yanı sıra, ortamın ph ından da etkilenir. Çünkü iyonik grupların dissosiyasyon derecesi ve polielektrolitin yük yoğunluğu ortamın ph ına bağlı olarak değişir. Polielektrolit çözeltilerinin diğer bir özelliği ise yüksek iyonik iletkenlikleridir. Elektrik alanı altında, küçük molekül ağırlıklı iyonlardan daha yavaş olmakla birlikte makroiyonlar da göç etmektedirler li yılların başlarında immünologların ve kimyacıların ortak çalışmaları ile bazı sentetik polielektrolitlerin organizmadaki immün cevaba etkisi aydınlatılmış ve sentetik polielektrolitlerin bu gibi amaçlar için daha uygun olduğu tespit edilmiştir. Çünkü polielektrolitlerin sentezi ve modifikasyonu daha basittir, istenilen molekül ağırlığında, elektrik yükünde, konformasyonda veya yüksek moleküler yapıda elde etmek mümkündür. Ayrıca suda iyi çözünür ve bilinen yapılarda çeşitli kompleksleri sentezlenebilir. Polielektrolitlerin molekül ağırlığı, polimerleşme derecesi ile orantılı olup molekül ağırlığı çözünürlüğü etkiler (Mustafaev, 1996) Homopolielektrolitler Homopolielektrolitler bütün monomerlerinde aynı işaretli yüklere sahip olan polielektrolitlerdir. Yüklü grupların tümünün eksi yüklü olması durumunda polianyon adını alırken, tümü artı yüklü ise polikatyon adını alır. Polielektrolitler, herbir tekrarlayan birimi bir elektrolit grup taşıyan polimerlerdir. Bu elektrolit gruplar sulu çözeltilerde dissosiye olup polimeri yüklü hale getirirler (Şekil 4.2). Polielektrolitlerin özellikleri hem elektrolitlere (tuzlar) hem de polimerlere benzemektedir ve bazen polituzlar olarak adlandırılmaktadırlar. Sulu çözeltileri, tuzlar gibi elektriği iletir ve polimerler gibi viskozdur. Şekil 4.2 Poliakrilik asitin sulu çözeltideki dissosiyasyonu Poliamfolitler Poliamfolitler yapılarında kovalent bağlı olarak anyonik ve katyonik grupların her ikisini de içeren makromoleküllerdir. Doğada bol bulunan veya sentetik yapıda elde edilebilen
39 25 proteinler, asidik veya bazik aminoasitler içerdikleri için amfolit özelliktedir. Bu proteinler ph a bağlı olarak proton verme veya alma özelliğindedir. Asidik ortamlarda proteinler çözeltiden proton aldıkları için net pozitif bir yüke, bazik ortam koşullarında ise ortama proton verdikleri için net negatif yüke sahip olurlar. İzoelektrik noktada ise poliamfolitlerin net yük toplamı sıfıra eşittir (Higgs ve Joanny, 1991; Merle,1987). Poliamfolitlerin yapısal özellikleri, anyonik ve katyonik monomer birimleri arasındaki iyonik çekme kuvvetleri tarafından yönlendirilmektedir. Anyonik ve katyonik grupların molar oranı birbirine yaklaşmaya başladıkça iyonik etkileşimler globular yapıya neden olurlar ve birçok durumda deiyonize suda çözünmezler (Higgs, 1991; Merle,1987). Bir poliamfolit izoelektrik noktasının ph koşullarında bulunuyorsa çözünürlüğü düşüktür ve kolay çöktürülebilir (Gözükara, 1997). 4.3 Polimerlerin Tıp ve Biyoteknolojide Kullanımı Geçen 30 yıl boyunca polimerlerin biyotıp alanında rolü önemli ölçüde artmıştır. Polimerik materyaller tarafından sağlanan fiziko kimyasal özellikler, biyomedikal alanda birçok kullanım alanına sahip olmasını sağlamıştır (Çizelge 4.1). Yeni yapısal mimari ile fonksiyonel polimerlerin geniş bir aralığı biyolojik çevrede geliştirilmiştir (Davis vd., 1978; Duncan, 2003). İlaç dağıtım çalışmalarında, yapay organ ve doku mühendisliğinde, stent gibi medikal malzemelerin hazırlanmasında, kontak lenslerde, dental malzeme üretimlerinde polimerik materyaller kullanılmaktadır (Kope cek vd., 2000). Çizelge 4.1 Bazı biyomedikal polimerlerin önemli kullanım alanları (Akovalı, 1995). Polimer Tipi Silikonlar Polietilen Polimetil Metakrilat Polivinil klorür Kullanım Alanları Yumuşak doku (göğüs, yanak vb.), kan ve sıvı boruları, kalp destek cihazları, implant kaplamaları, yarı geçirgen ve kontakt lensler, kontrollü ilaç salınımı Eklemler Sert kontakt lensler, intraoküler lensler, diş dolguları, kemik yapıştırıcıları Tıbbi borular, sıvı ve kan torbaları
40 26 Polietilen tereftalat Poliüretanlar Polikarbonat Polihidroksimetil metakrilat Polivinil prolidon Polisiyano akrilatlar Selüloz türevleri Kan damarları, kalp kapakçık halkası örtüsü Kalp destek cihazları, kanla temas yüzeyleri Yarı geçirgen sert kontakt lensler, yapay kalp bölmeleri Yumuşak kontakt lensler, yanık örtüleri, biyoelektrodlar, kontrollü ilaç salınımı Kan hacmini artırıcı, yumuşak kontakt lensler Yumuşak ve sert doku yapıştırıcıları Yarı geçirgen kontakt lensler, membranlar, kontrollü ilaç salınımı Polimerler esas olarak biyoaktif olabilir veya kimyasal ve biyoterapötik ajanlar olarak çeşitli alanlarda spesifik olarak kullanılabilir. Ringsdorf un sunduğu orijinal çalışmada, bölge hedefli polimerik ilaçlar, çeşitli hastalıklara karşı terapötik ajanlar olan fonksiyonel polimerlerin rollerini aydınlatmıştır. Bu polimer ilaç-konjugat sistemleri küçük moleküllü ilaçların ve terapötik ajanların hedefli dağıtımını sağlamıştır (Şekil 4.3) (Harris ve Chess, 2003). Şekil 4.3 Polimer-ilaç, polimer-protein ve polimer-dna konjugat sistemleri (Abe vd., 2006). Son zamanlarda birçok fonksiyonel polimer, gen terapide genetik materyalin dağıtımı için non-viral vektörler olarak geliştirilmektedir (Nucci vd., 1991; Russell-Jones, 1996). Polimerilaç konjugatları gibi polimer bazlı ilaç geliştirme alanındaki çabalar, klinik olarak
41 27 uygulanabilen ürünleri içererek bu alanda kayda değer bir ilerleme sağlamıştır (Okamoto, 1998; Takakura vd., 1998; Allen, 2002). Başlangıçta yenilikçi polimer temelli ilaçlara karşı konulmuştur ancak bilimsel meraklar, endüstriyel gelişmeler sonucu düzenleyici birimlerin onayları da alınarak bazı gereksinimler için polimer uygulamaları çalışılmıştır. Klinikte düşük maliyet ile verimli ilaç ve tanı sistemleri olarak güvenli şekilde uygulanması amaçlanmıştır. Polimer tedavisi, klinik amaçlı nanoteknoloji olarak adlandırılan popüler alanda odak noktası olup bu alanda büyük bir gelişime neden olmuştur (Abe vd., 2006). 4.4 Poliakrilik Asit Poliakrilik asit, akrilik asit monomerlerinden oluşan bir polimerdir. Akrilik asit oda sıcaklığında renksiz ve kokulu bir sıvıdır. Su, alkol, eter ve birçok organik çözücü ile çözünebilen bir maddedir. Akrilik asit ısıya, ışığa veya metallere maruz bırakıldığında çok kolay polimerleşir. Şekil 2.2 de görüldüğü gibi akrilik asitin kimyasal yapısı CH 2 =CH- COOH dır (Willey, 1964). Şekil 4.4 Poliakrilik asitin yapısı Akrilik asitin bazı özellikleri Çizelge 4.2 de verilmiştir (Willey, 1964). Akrilik asitin kolaylıkla polimerize olma özelliğinden dolayı kuvvetli bir ekzotermik polimerleşmeden korunmak için ticari akrilik asitin içerisine polimerleşme inhibitörü eklenir. Poliakrilik asitin çözünürlüğünü etkileyen birçok parametre vardır. Bunlara deney şartları, dallanma derecesi, molekül ağırlığı, çapraz bağlanma oranı, ortamdaki diğer çözünen maddeler ve sıcaklık örnek verilebilir (Barreiro-Iglesias vd., 2004). Poliakrilik asitin kurutma işlemi çapraz bağlanmayı önleyecek şekilde yapılırsa çözünürlüğü yükselir. Buna karşın polimer yüksek sıcaklıklarda tamamen kurutulmuşsa çözünürlüğü oldukça azdır (Willey, 1964).
42 28 Çizelge 4.2 Akrilik asitin bazı özellikleri (Willey, 1964) Molekül ağırlığı (g/mol) 72,0 Donma noktası ( o C) 13,5 Yoğunluk (g/ml) 1,045 (25,0 o C ) Normal kaynama noktası ( o C) 141,0 Refraktif indeksi 1,4185 (25,0 o C ) Kinematik viskozitesi (cks) 1,1 (25,0 o C) Poliakrilik asitin çözünürlüğü sıcaklığın azalmasıyla düşer. Fakat poliakrilik asitin molekül ağırlığı ve konsantrasyonu düşükse ve tuz ya da mineral asitleri bulunduruyorsa sıcaklık düştüğünde çözünürlüğünde azalma görülmez. Ancak hidrojen bağı olan bileşiklerdeki çözünürlüğü sıcaklığın artması ile düşer (Willey, 1964). Akrilik asit farklı alanlarda kullanılmaktadır. Örneğin tekstil materyallerinin modifiye edilmesinde, plastiklerde, kâğıt imalatında, dış cephe boyalarında, zemin cilalarında, zemin ve duvar kaplamalarında, otomobil, araç-gereç ve mobilya verniklemede, eczacılıkta kontrollü ilaç salınım sistemlerinde kullanılmaktadır. Biyo-adeziv hidrojel olarak, hem mukozaya yapışan hem de viskozite artıran PAA jelleri, gözde merhemler gibi bulanıklık yapmadan rahat şekilde kullanılmaktadır (Robinson, 1989). Göz için yapılan ilaçların, göz içine iyi bir şekilde absorplanabilmeleri ve tedavide aktif rol oynayan maddelerin daha uzun süreli göz tabakasında bulunabilmeleri için PAA ile deneyler yapılmıştır. Yapılan bir çalışmada PAA gibi ayrı ayrı kullanılan polimerler mukoz tabakayla bağ kurarak göz yüzeyinde çözünür, kolloidal ve partikül halde bulunan materyali yakalayabilmektedir. Vajinal lezyonların tedavisi amacıyla poliakrilik asit, metilselüloz, karboksi metilselüloz, hidroksi propil selüloz ve hidroksipropil metil selüloz kullanılarak asiklovir içeren bioadhesive tabletler hazırlanmıştır. Bioadhesive polimer olarak poliakrilik asit ve hidroksipropil metil selüloz kullanılan tabletlerin laktik çözeltisi veya distile suda 6 saat boyunca şiştiği ve etkin madde salımının 8 saatten daha fazla olduğu görülmüştür. Poliakrilik asit, sodyum karboksimetil selüloz ve çinko sülfat içeren vajinal tabletler bu konuda önemli örneklerdir (Bell vd., 1995; Serres vd., 1996; Ramkinssoon vd., 1999; Genç vd., 2000).
43 29 Yapılan bazı çalışmalarda poliakrilik asitin adjuvant, antitrombojenik madde ve ilaç salınımlarında taşıyıcı olarak kullanılmakta olduğu gösterilmiştir. Normalde toksik olan PAA zincirine, NIPAAm ünitelerinin katılmasıyla toksisitesi azaltılmaktadır (Mustafaev vd., 1998). Mevcut çalışmalar, poliakrilik asit ve Cu 2+ komplekslerinin sulu çözeltilerinin, protein karışımları ile radyasyon etkisine maruz bırakıldıklarında, belli dozlara kadar gerek polimer gerekse protein moleküllerinin in vitro koşullarda bozulmadan direnç gösterdiklerini bildirmiştir. Poliakrilik asit ve poliakrilik asit Cu 2+ örneklerinin kan hücrelerini radyasyondan koruma özelliği taşıdığı gösterilmiştir (Mustafaev vd., 1998). 4.5 Biyouyumluluk ve Toksisite Testleri Biyouyumluluk kavramı; spesifik uygulama için kullanılacak materyalin, toksik olmaması ve fonksiyonel şekilde çalışması olarak tanımlanmıştır. Son 10 yıldır tartışılan biyouyumluluk kavramında genel olarak iki prensip vardır. Bunlardan birincisi sitotoksik etkinin saptanması, ikincisi ise biyofonksiyonelliğin gösterilmesidir. Biyomateryal veya ilaç sistemlerinin fonksiyonelliğini belirlemek için kullanım amacına göre testler seçilir. Ancak ne kadar fonksiyonel olursa olsun, insanda kullanılması hedeflenen maddelere klinik öncesi toksisite testlerinin yapılması şarttır (Kirkpatrick vd., 2005). Toksisite testleri in vivo hayvan modellerinde ve in vitro kültür modellerinde yapılmaktadır. Deney hayvanlarını koruma yasalarının temelindeki 3R (reduction, replacement and refinement) kuralına göre; toksisite çalışmalarında deney hayvanlarının kullanılması yerine sayısı hızla artan yeni alternatif yollar geliştirilmektedir. Bunlardan biri olan in vitro toksikoloji yöntemleri kullanılarak yapılan hücre kültür çalışmaları, son yıllarda yaygınlaşmış ve laboratuar hayvanı kullanımı yerine daha ekonomik olan alternatif bir yol getirmiştir. İn vitro toksikolojik teknikler pratik, güvenilir ve avantajlı yöntemler olarak bilinmektedir. Bu teknikler ürünlerin geliştirilmesi, ilaçların keşfi, güvenliğin geliştirilmesi ve in vivo akut ölümcüllüğün önceden anlaşılmasında rutin olarak kullanılmaktadır. Birçok çalışmada insanda akut toksisite ile in vitro metotlar arasında korelasyon olduğu gösterilmiştir (Lestari vd., 2006). Ancak hücre kültürü asla tam bir organizmanın cevabını oluşturmaz. Organizmada meydana gelebilecek etkileşimler hakkında tam olarak bilgi vermez. Diğer bir problem de memeli kültürünün en çok kullanılan kalıcı hücre serileri, biyomateryalin toksik etkilerine primer kültürlerden veya birkaç pasajlık kültürlerden daha az duyarlıdır. Ayrıca devamlı veya primer hücre serileri arasındaki seçime ek olarak, hücre türünün seçimi de oldukça önemlidir.
44 30 Uygun testin dizaynında uygulanan daha ileri bir prensip insandan alınan biyolojik dokulardır. İlk olarak izole edilen doku veya bağlantılı hücreler taze izole edilmiş primer yapıda olup transforme olmamalıdır. Fakat primer kültürlerin insandan izole edilmesi ve kültürünün yapılması oldukça zordur. Primer kültürler farklı donörlerden alındığı için fonksiyonel durumları yansıtması farklıdır. Kalıcı hücre serileri bu açıdan daha stabil bir fenotipe sahiptir. Ayrıca kalıcı hücre serileri ile yapılan çalışmalar, primer hücrelere göre tekrar edilebilirdir. Kalıcı hücrelerin; kendiliğinden, viral antijenlerle veya kimyasallarla transforme olması bunların önemli hücresel fonksiyonlarını denetlemeyi gerektirir. İn vitro kültür metotlarında diğer bir önemli durum ise kültürde yapının 2 boyutlu olmasıdır. Geçerli fonksiyonel parametreleri içermesi için gerekli hücre fenotipinin sağlanması oldukça önemlidir. Bir hücre tipinin saf izolasyonunun sağlaması yeterli değildir. Ayrıca kültür koşullarında gerekli hücresel fonksiyonlarının kaybolma riskinin olmadığı gösterilmelidir (Kirkpatrick vd., 2005). Uygun hücre kültürü seçimi ile beraber kullanılacak toksisite testinin seçilmesi de oldukça önemlidir. İn vitro sitoksisite testlerinde; kültür ortamında hücreler ile belli süre etkileşime bırakılan maddelerin hücrelerde oluşturduğu değişimler ve hücre canlılığı incelenir. Hücrelerin metabolik aktivitesinin ölçülmesine dayalı birçok kolorimetrik yöntem geliştirilmiştir. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) veya 3- (4,5-dimetiltiazol-2-il-)-5-(3-karboksimetilmethoksifenil)-2-(-4-sulfofenil)-2H-tetrazolyum (MTS) gibi testler ile hücre canlılığı ölçülebilir. Hücre sayısının belirlenmesinde diğer bir kolorimetrik metot ise toplam DNA miktarının ölçülmesidir. Kullanılan floresan boya çift sarmal DNA nın nükleik asitlerine bağlandıkça daha yoğun floresan açığa çıkar. DNA miktarının ölçülmesinde yüzeye tutunan hücrelerin dezavantajı hücrelerin tripsinizasyonunu gerektirmesidir. Sonuçta materyalden kaldırılması ve boyamayı gerektirmesi zahmetli ve yoğun bir süreçtir. Radyo etiketli timidin ile çoğalan hücre sayısı tam olarak ölçülebilir. Radyoaktif timidin ekleme yöntemiyle yeni bölünen hücrelerin DNA molekülleri işaretlenir. Geriye kalan radyoaktif timidinin ortamdan uzaklaştırılması ve örneğin radyoaktivitesinin ölçümü artan hücre sayısı ile doğrudan orantı gösterir. Bu metot başlangıç hücreleri radyoaktif timidini kullanmadıkları için bütün hücre sayısını ölçemez. Diğer bir dezavantajı ise radyoaktivite kullanmanın neden olabileceği sağlık riskleridir. Radyoaktivitenin getirdiği problemler, timidin analoğu olarak 5-bromo-2 -deoksiüridin kullanılarak uygulanmasıyla ortadan kaldırılabilir.
45 31 Hücre fenotip ve canlılığı ile yakın ilişkili olduğundan, hücre morfolojisinin görüntülenmesi önemlidir. Floresein diasetat veya klorometilfloresein diasetat boyalar hücre fiksasyonunu gerektirmeden canlı hücrelerin klasik ışık mikroskobu ve elektron mikroskobu kullanılmasıyla gözlenmesini sağlar. Floresan olmayan lipofilik boyalar, hücre membranından içeri alınıp hücresel hidrolaz enzimi ile parçalanarak hücre dışına kolayca çıkamayacak olan floresan ürünler oluşturur. Hücre morfolojisi floresan mikroskop veya konfokal lazer taramalı mikroskop ile kolayca gözlenebilir. Bu boyalar hücre büyümesine negatif bir etki yaratmadığından dolayı uzun süreli kültürü gözleyebilmek amacıyla kullanılabilir. Atomik force mikroskobu gerçek zamanlı dinamik hücresel sürecin çalışılması için potansiyel bir araç olabilmektedir. Fiziksel tampon aday hücrelerin morfolojiyi bozmadan ve hasara yol açmadan devamlı görüntülenmesi amacıyla hücrelere saatlerce uygulanabilir. Hücrelerin yayılımı, hareketliliği gibi dinamik olaylar normal kültür sırasında gözlenebilir. İmmunofloresan; hücre yapısının tanımlanmasında, hücrelerin dinamik proseslerinin, nükleus, lizozom, sitoiskelet gibi elemanların görüntülenmesinde kullanılan önemli bir metottur. Direkt immunofloresan yöntemi ile komponentler hücresel hedef moleküllerce tanınır ve doğrudan floresan boyayla boyanır. İndirek immunofloresanda ise sandviç metodu ile çalışılır. Antikor ilk olarak hücresel hedef moleküle (antijene) bağlanır, ikinci adımda antijen-antikor kompleksi boya bağlı antikorlar ile belirlenir. Canlı hücrelerin nükleusunu boyamada Hoechst 33342, 4-,6-d diamidino-2-fenilindole, dihidroklorid (DAPI) ve etidyum bromid rutin olarak canlı hücrelerin nükleusunu boyamada kullanılır (Ma vd., 2007). Hücrelerin biyosentetik fonksiyonlarının incelenmesindeki genel prensipler kısaca şöyledir. İlk olarak in vitro hücre fenotipinin başlıca elemanlarının incelenmesi esastır. Genelde bu, insan primer hücrelerinin erken pasajda kullanılması anlamına gelir. İkinci olarak hücre biyosentetik fonksiyon kinetiğinin çeşitli yönleri araştırılmalıdır. Bu özellikle doz-cevap çalışmalarında farklı zaman noktalarının kullanımını içerir. Doz-cevap deneyleri adezyonu artıran mikromolar altı iyon konsantrasyonu ile ilgilidir. Bu in vitro deneylerde hücreleri baskılar, organizmaya girişte dikkatli karar verilmesini gerektirir. Üçüncü olarak biyosentetik aktivite sadece gen üretim seviyesinde değil, transkripsiyon seviyesinde de önemlidir. Gen ekspresyonunun her iki seviyesinin çalışılması hücre içinde gen kontrol ve sinyal üretiminin anlaşılması için gereklidir (Kirkpatrick vd., 2005). Biyomateryallerin sebep olduğu sistemik toksisite ve mekanik hasarın belirlenmesinde in vitro kültür modelleri kullanılmaktadır ancak hayvan denemelerinde olduğu kadar net sonuçlara ulaşılamaz. İn vitro testlerin sistemik toksisiteyi araştırmakta yetersiz kalabileceği,
46 32 bu nedenle ancak in vivo testler için bir ön tarama testi olarak kabul edilmesi gerektiği şeklinde tartışmalar sürmektedir (Davis, 1994). Bu kültür metotları ile maddelerin vücutta oluşturacağı etkileşimler tam olarak ortaya konulamayacağı düşünülmektedir. Ancak kültür yöntemi ile polimerlerin sitotoksik özellikleri ve yıkım ürünleri tanımlanabilir. Bunun yanısıra in vitro deri absorbsiyon testi, in vitro fototoksisite testi, in vitro deri korozyon testi ve hassasiyet testleri hayvan testlerinin yerini almış testlerdir. Kimyasalların, ilaçların, kozmetik ürünlerin toksisite testlerinin şu anki ve gelecekteki düzenlemeleri için in vitro metotların ve hücre kültürü yöntemlerinin geliştirilmesi gereklidir (Lestari vd., 2006) (4,5-dimetiltiazol-2-il)-2,5-difeniltetrazolyum (MTT) Testi Hücreler üzerindeki toksik etkinin incelenmesi amacıyla hücre canlılığının ölçülmesini sağlayan metotlardan önemli bir tanesi de MTT testidir. MTT testi hücre büyümesi veya hücre ölümünü değerlendirmeye olanak sağlayan indirekt bir testtir. Kültür ortamındaki hücreler üzerinde uygulanabilen ve hücrelerin metabolik aktivitesinin ölçülmesine dayalı kolorimetrik bir yöntemdir. Hücrelerin mitokondrilerinde bulunan dehidrojenaz enzimi ile MTT reaktanı mor renkli formazan kristaline indirgenir. Bu reaksiyon sadece canlı hücrelerde gerçekleşir. Oluşan kristaller, mikroskobik olarak gözlemlenebilir ve DMSO, izopropanol gibi kimyasallar ile çözündürülerek oluşan mor renk kolorimetrik olarak ölçülebilir. Mor renkli ürünün toplam miktarı hücre popülasyonundaki dehidrojenaz enziminin toplam aktivitesini verir. Ölçülen optik değer hücrelerin canlılığı ile doğrudan orantılıdır (Kirkpatrick vd., 2005).
47 33 5. DENEYSEL ÇALIŞMALAR 5.1 Deneysel Çalışmalarda Kullanılan Cihazlar ve Kimyasal Maddeler Deneysel Çalışmalarda Kullanılan Cihazlar Deneysel çalışmalarımızda aşağıdaki cihazlar kullanıldı. Laminar-hava akışlı kabin Soğutmalı Etüv (Friocell) İnkübatör (37 C %5 CO2) Ters mikroskop ELISA okuyucu Magnetik karıştırıcı (Heidolp MR 3001) ph metre (İnolab WTW level 1) Hassas terazi (Precisa XB 220A) Kaba terazi (Precisa BJ 6100D) Santrifüj (Hettich Zentrifugen-EBA20) Vorteks (Heidolp REAX top) Bidistile su cihazı Buzdolabı -45 C derin dondurucu Sıvı azot tankı Otoklav Flow Sitometri (Beckman Coulter- Cell Lab Quanta)
48 Deneysel Çalışmalarda Kullanılan Kimyasal Maddeler ve Besiyerleri MADDE ADI ÜRETİCİ FİRMA KATALOG NO Sodyum hidroksit Riedel De Haen Hidroklorik asit Merck RPMI 1640 with L glutamin Biological Industries B L-glutamin Biological Industries IC Penisilin streptomisin Biological Industries IC Gentamicin Sigma G1397 Fetal sığır serumu Seromed S0115 Tripsin EDTA Biological Industries B DMSO Sigma D26650 Sodyum klorür Atabay AT Potasyum klorür Carlo Erba Disodyum hidrojen fosfat Riedel De Haen Potasyum dihidrojen fosfat Riedel De Haen 8210A Tripsin Biochrom L2103 EDTA Merck Thiazoly blue tetrazolium bromide(mtt) Sigma M5655-1G Formaldehyd (%37) Applichem A propanol Applichem A03928 Sodyum dodesil sülfat (SDS) Merck S Poliakrilik Asit (MA:100,000 ) Aldrich Poliakrilik Asit (MA:30,000 ) Aldrich Annexin-V Invitrogen A AAD Invitrogen A
49 Çözeltilerin Hazırlanması x PBS Tampon Hazırlanması 8 g NaCl, 0,2 g KCl, 1,44 gr Na2HPO4, 0,24 g KH2PO4 tartılıp 900 ml deiyonize suda çözündürüldü. ph 7,4 e ayarlandı ve son hacim deiyonize suyla 1 litreye tamamlandı. 1 atm basınç, 121 C sıcaklık ayarlanmış otoklavda cam şişeler içersinde 20 dakika steril edildi x Tripsin solusyonu Tripsin 62,5 mg tartıldı. Steril edilmiş 50 ml lik 1xPBS içinde çözündürüldü. Önce 0,45 lik sonra 0,22 lik filtreden süzülerek steril edildi RPMI 1640 Medyumunun Hazırlanması 100 ml RPMI 1640 (L-glutamin içeren) içerisine 0,5 ml gentamisin (10 mg/ml lik) eklenerek stok hazırlandı. Kültürde kullanılmak üzere buradan 45 ml alınıp 5 ml FCS eklenerek %10 FCS li besiyeri hazırlandı Hücre Dondurma Medyumunun Hazırlanması %80 FCS ve %20 DMSO dan steril bir biçimde hazırlandı MTT Solüsyonunun Hazırlanması MTT (Sigma M5655) 1xPBS içersinde çözündürüldü. Kullanılacak MTT prosedürüne uygun olarak 0,6 mg/ml ve 10 mg/ml konsantrasyonlarda 5 ml hazırlandı. Önce hassas terazide 0,6 mg/ml için 3 mg, 10 mg/ml için 50 mg MTT tartılıp PBS ile 5 ml ye tamamlandı. Vortekste çalkalanarak iyice çözündükten sonra steril edilmek üzere önce 0, 45µl daha sonra 0,22 µl lik (CS) filtreden geçirilerek steril edildi. MTT ışığa duyarlı olduğu için, genel olarak bu işlemler karanlıkta yapıldı MTT Reaksiyon Sonlandırıcı Solüsyonun Hazırlanması %50 izopropanol, %10 SDS bulunan bu çözeltiden 10 ml hazırlandı. 5 ml sıvı izopropanol, 5 ml distile su ve 1 mg toz SDS tartılıp vorteksle karıştırılarak hazırlandı.
50 Poliakrilik Asit (50 mg/ml, MA: 100,000 g/mol) Çözeltisinin Hazırlanması Deneylerde MA: olan ve ticari olarak satın alınan %35 g/ml sıvı PAA kullanıldı. PAA %35 lik olduğu için 100 ml de 35 g PAA bulunur. 50 mg dan 5 ml hazırlamak için 2500 mg PAA gereklidir. 100 ml de 35 g varsa X 2500 mg X=7,142 ml PAA 35 ml PBS eklendi saat manyetik karıştırıcıda karışması beklendi. ph 7 ye ayarlandı. Son hacim 50 ml olacak şekilde PBS ile tamamlandı, 0,22 µm lik millipore filtrelerden geçirilerek steril edildi ve +4 C de saklandı Poliakrilik Asit (50 mg/ml, MA: 30,000) Çözeltisinin Hazırlanması MA: 30,000 olan ve ticari olarak satın alınan %40 g/ml sıvı PAA kullanıldı. 100 ml sinde 40 g PAA bulunacağı için 100 ml 40 g varsa X 2500 mg X= 6,25 ml PAA PBS eklenip manyetik karıştırıcıda 48 saat karıştırıldıktan sonra ph 7 ye ayarlanıp son hacim 50 ml olacak şekilde PBS ile tamamlandı. 0,22 µm lik millipore filtrelerden geçirilerek steril edildi ve +4 C de saklandı Bağlayıcı Tampon (Binding Buffer) Çözeltisi Hazırlanması 50 ml distile su içinde son konsantrasyon 10 mm HEPES/NaOH, 150 mm NaCl, 5 mm KCl, 1 mm MgCl 2 ve 1.8 mm CaCl 2 olacak şekilde ph 7.4 e ayarlandıktan sonra +4 C de saklandı Annexin V-FITC Solüsyonu Hazırlanması 1 mg/ml olan Annexin V stok çözeltisinden 10 µl alınıp 90 µl bağlayıcı tampona (Binding Buffer) eklenir. 100 µl çalışma solusyonundan her örnek için 10 µl kullanılır.
51 AAD Çalışma Solüsyonu Hazırlanması 1 mg/ml olan 7-AAD (Sigma) stok çözeltisinden 20 µl alınıp 980 µl PBS ye eklenir µl çalışma solusyonundan her örnek için 5 µl kullanılır. 5.2 Hücre Kültürü Teknikleri Hücrelerin Kriyobanktan Çıkarılması J774 Hücre Hattının Kriyobanktan Çıkarılması Sıvı azot tankından çıkartılan hücreler 37 C su banyosunda çalkalanarak sıvı içersinde tutularak çözündürüldü. Çözünen hücre süspansiyonu tüpüne alınarak üzerine 5 ml %10 FCS li RPMI 1640 eklenir ve 1000 rpm de 5 dakika santrifüj edildi. Süpernatant döküldükten sonra pellete az miktar besiyeri eklenerek yavaşça pipetaj yapıldı ve besiyeri konulmuş flasklara ekim yapıldı MONI/EP126 L. infantum Suşlarının Kriyobanktan Çıkarılması Sıvı azot tankından çıkartılan hücreler 37 C su banyosunda çalkalanarak sıvı içersinde tutularak çözündürüldü. Çözündürülen hücre süspansiyonunun, 7,5 ml RPMI 1640 (%10 FCS) besiyeri konulmuş flasklara ekimi yapıldı Hücre Hatlarının Kültürü J774 Hücre Hattının Kültürü Bu çalışmada kullanılan J774 serisi makrofajlar, İstanbul Üniversitesi Tıp Fakültesi Histoloji ve Embriyoloji Anabilim dalı öğretim üyesi Sayın Prof. Dr. Ayhan Bilir tarafından temin edilmiştir. Hücreler, Yıldız Teknik Üniversitesi, Hücre Kültürü Laboratuvarı na, devamlı kültürleri yapılmak üzere getirildi ve kültürün adaptasyonu sağlandı. Kültür labotuvara getirildikten sonra kendi besiyerinde tutuldu. 48 saat sonra ise kültür ortamında bulunan 5 ml besiyerinin 2,5 ml si alınarak yerine yeni hazırladığımız besiyerinden (RPMI %10 FCS) aynı miktarda eklendi. Kültürün durumu her gün ters mikroskop ile incelendi. Besiyeri, kültürün durumuna ve ortamın ph sına uygun olarak değiştirildi. Yüzey kaplamasını tamamladığı tespit edilen makrofajların pasajı yapıldı. Pasajın yapılması için hücrelerin devamlı kültürü %10 FBS içeren RPMI 1640 besiyerinde yapıldı. Kültür kabı olarak 25
52 38 cm 2 lik steril kültür flaskları kullanıldı. Kültür %5 CO 2 içeren etüvde (Innova CO-150) 37 C de inkübasyona bırakıldı. Ters mikroskop ile günlük gelişimleri ve hareketlilikleri incelendi. Genel olarak pasajı haftada bir kez yapıldı. Yüzeyi kaplayan hücreler tripsinizasyon ile kaldırıldı. Tripsinizasyon işlemi için önce flask içerisindeki besiyeri döküldü ve PBS ile 2 defa yıkandı. 2 ml Tripsin-EDTA eklenerek 37 C de 5 dakika inkübe edildi. Hücreler yüzeyden ayrıldıktan sonra 7 ml RPMI 1640 (%10 FBS) içeren besiyeri içerisinde 1000 rpm de 5 dakika santrifüj edildi. Süpernatant dökülerek dipteki hücre pelletinden makrofajların sayımı yapılarak ml deki makrofaj sayısı hesaplandı. Flask içindeki 5 ml besiyerinde 5x10 5 makrofaj olacak şekilde ekim yapıldı. Ayrıca pasaj sırasında elde edilen kültürün geri kalan kısmı kriyoprezervasyonu yapılarak kriyobankın oluşturulması için kullanıldı Tripan Mavisi (TM) ile J774 Hücre Canlılığının Tayin Edilmesi ve Hücre Sayımı Tripan mavisi (Biol. Ind., ) molekül yapısı C 34 H 23 N 6 O 14 S 4 Na 4 olan, canlı hücreleri boyamayıp ölü hücreleri boyayan böylece canlı hücre sayısını tespit etmekte kullanılan vital bir boyadır. Hücrelerin pasaj ve deneylerde kullanılmadan önce sayımı, tripan mavisi ile thoma lamında yapıldı. Sayımın kolay yapılması için 50 kat seyreltme tercih edildi. Kültür flaskı içerisindeki besiyeri döküldü ve PBS ile 2 defa yıkandı. 2 ml Tripsin-EDTA eklenerek 37 C de 5 dakika inkübe edildi. Hücreler yüzeyden ayrıldıktan sonra 7 ml RPMI 1640 (%10 FBS) içeren besiyeri içerisinde 1000 rpm de 5 dakika santrifüj edildi. Süpernatant dökülerek dipteki hücre pelletinden makrofajların sayımı yapılarak ml deki makrofaj sayısı hesaplandı. Hücre süspansiyonundan 2 µl alınarak 1,5 ml lik steril eppendorf tüpü içine konuldu. Bu oranın sağlanması için üzerine 48 µl tripan mavisi solüsyonu ve 50 µl besiyeri eklenerek iyice pipetaj yapıldı. Thoma lamının her iki bölümüne de lam ile lamel arasına 10 ar µl hücre süspansiyonundan dikkatlice yayıldı. Ters mikroskopta tripan mavisi ile maviye boyanmış ve boyanmamış olan hücreler sayıldı ve kültürün % canlılık oranı hesaplandı. Hücre canlılığı %95 in altında olan kültürler deneylerde kullanılmadı.
53 MONI/EP126 Leishmania Promastigot Kültürünün Yapılması VL etkeni olan L.infantum promastigotları (MONI/EP126), Ege Üniversitesi Tıp Fakültesi Parazitoloji Anabilim Dalı öğretim üyesi Sayın Prof. Dr. Seray Özensoy ve Prof. Dr. Yusuf Özbel den temin edildi. Leishmania parazitlerinin kültürü RPMI 1640 besiyerlerinde 27 C soğutmalı etüvde (Friocell, 111) gerçekleştirildi. Parazitlerin gelişimi ve morfolojik durumu ters (invert) mikroskopta (Olympus CKX41) günlük olarak incelendi. Kültürün pasajı haftada bir kez yapıldı. Pasaj öncesi kültürden örnek alındı, ortamın ph ı parazitlerin morfolojileri, hareketlilikleri incelendi ve sayımları yapıldı. Flaska 7,5 ml besiyeri eklendi ve üzerine 3x10 5 parazit/ml olacak şekilde kültürden parazit ekimi yapıldı. Leishmania promastigotlarının kültürü bu şekilde devam ettirildi Parazitlerin Thoma Lamında Sayımı Kültür ortamındaki parazitler hareketli oldukları için thoma lamında sayılması zordur. Bu nedenle homojenliği sağlanmış kültürden ~ µl alındı ve 1:1 oranında %2 lik formalinle (sigma, HT501128) karıştırılarak 5 10 dakika bekletildi. Daha sonra Thoma lamında (Menzel superior) aşağıdaki formül kullanılarak sayım yapıldı: Parazit sayısı = Ortalama hücre sayısı x Sulandırma katsayısı x Thoma lamı sabiti Parazit sayısı: 1 ml deki parazit sayısı Ortalama hücre sayısı: Thoma lamının alt ve üst kısmında bulunan 16 karedeki hücrelerin aritmetik ortalaması Sulandırma katsayısı: Sayım için yapılan sulandırma katsayısı Thoma lamı sabiti: Hücrelerin Dondurulması ve Saklanması J774 Hücrelerinin Dondurulması ve Saklanması Flask yüzeyini tek tabaka olarak kaplayan makrofaj kültürü üzerine Tripsin-EDTA solüsyonundan 2 ml. ilave edilerek hücrelerin ayrılması sağlandı. Derhal üzerine 7 ml kültür medyumu ilave edilerek hücreler pipetle steril santrifüj tüplerine aktarılıp 1000 rpm de 5 dakika, +4 C de santrifüj edildi. Santrifüj edilen tüplerin üst fazı döküldükten sonra dipte kalan pellet resüspanse edildi ve son hacim, %10 DMSO içeren dondurma solüsyonu ilave edilerek kriyo tüplere aktarıldı. Kriyo tüpler +4 C de 1 saat, 20 C de 2 saat kaldıktan
54 40 sonra, 40 C derin dondurucuda 24 saat bekletildi, kriyobankı oluşturulmak üzere sıvı nitrojen tankına konuldu MONI/EP126 Promastigotlarının Dondurulması ve Saklanması Kültürün genel durumunun iyi olduğu invert mikroskop ile incelendikten sonra pasaj aşamasına gelen hücreler donduruldu. Süspansiyon halindeki kültürden 1,8 ml kriyo tüpe alınıp, 0,2 ml steril DMSO yavaşça eklendi. 1 saat 4 C de, 24 saat -40 C de bekletilip sıvı azot tankına konuldu. 5.3 Polimerlerin Toksik Etkisinin MTT Yöntemi ile İncelenmesi Canlılığı ölçülecek hücreler düztabanlı 96 kuyuculu plaklara ekildi. Yüzey kaplaması tamamlandıktan veya yeterli yoğunluğa ulaştıktan sonra (24 veya 48 saat) ortama polimer ilave edildi. Madde ile 48 saat inkübasyonun ardından 10 mg/ml konsantrasyonda hazırlanan MTT solüsyonundan her bir kuyucuğa 10 µl eklendi ve 37 C de 4 saat inkübe edildi. Formazan kristalleri oluşumu invert mikroskopla tek tek incelendi. MTT durdurucu solüsyondan 100 µl eklendi 30 dakika oda sıcaklığında beklenip formazan kristallerinin çözündüğü kontrol edildi. Aynı işlemler polimer besiyeri etkileşimini kontrol etmek amacıyla, hücre bulunmayan sadece besiyeri ilave edilmiş 96 kuyucuklu plaklarda da yapıldı. Son olarak optik yoğunluğu ölçmek üzere 96 kuyucuklu plağın kapağı açılarak 570 nm dalga boyuna ayarlanmış ELIZA cihazına yerleştirildi ve cihazın optik ölçüm sonuçları bilgisayar ortamından alındı PAA nın J774 Hücrelerindeki 48 Saatlik Toksik Etkisinin MTT Yöntemi ile İncelenmesi J774 hücreleri ml de 100,000 olacak şekilde fenol red siz RPMI 1640 (%10 FCS) besiyeri içersinde deneyde kullanılacak miktarda hazırlandı. Hazırlanan hücre süspansiyonundan her kuyucuğa 100 µl (10,000 hücre/kuyucuk) eklendi. Kuyucuk sayısı polimerin her konsantrasyonu için 4 kuyucuk olacak şekilde hesaplandı. Hücrelerin dağılımının eşit olduğu mikroskop ile incelendikten sonra, 37 C lik CO 2 etüvüne kaldırıldı ve 24 saat inkübe edildi. İnkübasyondan 24 saat hücrelerin yüzeyi kapladığı ve morfolojik olarak normal olduğu tespit edildikten sonra polimerler son konsantrasyon 2,5 µg/ml, 5 µg/ml, 10 µg/ml, 25 µg/ml, 50 µg/ml, 100 µg/ml, 250 µg/ml, 500 µg/ml, 1000 µg/ml ve 2500 µg/ml olacak şekilde eklendi.. Polimerlerin hazırlanan 50 mg/ml stoktan kullanıldı.
55 41 Polimer eklendikten sonra %5 CO 2 içeren etüvde 37 C de 48 saat inkübasyona bırakıldı. MTT solüsyonunun hazırlanması bölümünde açıklandığı gibi hazırlanan 10 mg/ml MTT solüsyonundan her kuyucuğa 10 µl eklendi ve 37 C de 4 saat inkübe edildi. Formazan kristalleri oluşumu invert mikroskopla tek tek incelendi ve fotoğrafları çekildi. MTT durdurucu solüsyondan 100 µl eklendi, formazan kristallerinin tamamen çözüldüğü mikroskobik olarak incelendi ve 30 dakika oda sıcaklığında beklenip ELIZA cihazı ile 570 nm dalga boyunda optik yoğunluk ölçüldü PAA nın MONI/EP126 Promastigotlarına 48 Saatlik Toksik Etkisinin MTT Yöntemi ile İncelenmesi Flaskta kültürü yapılan parazitler yeterince çoğaldıkları ve durumlarının iyi olduğu invert mikroskopta incelendikten sonra, hücre sayımı yapıldı. Süspanse kültürden ml de 300,000 parazit olacak şekilde mikropipet ile çekildi ve fenol red siz RPMI 1640 (%10 FCS) besiyeri içersinde dilüe edilerek deneyde kullanılacak miktarda hazırlandı. Hazırlanan hücre süspansiyonundan her kuyucuğa 100 µl (30,000 hücre/kuyucuk) eklendi. Polimerin her konsantrasyonu için 4 kuyucuk olacak şekilde hesaplandı. Hücrelerin dağılımının eşit olduğu mikroskop ile incelendikten sonra, 27 C lik soğutmalı etüvde 48 saat inkübe edildi. İnkübasyon sonunda hücrelerin genel durumlarının normal, hareketlerinin aktif olduğu tespit edildi ve polimerler son konsantrasyon 1, 5, 15 mg/ml olacak şekilde eklendi. Polimerlerin hazırlanan 50 mg/ml lik stoktan kullanıldı ve kuyucuklara aşağıdaki şekillerde eklendi: Kontrol grubu olarak hücresiz besiyeri üzerine aynı konsantrasyonlardaki polimerler aynı şekilde ilave edildi. Polimer eklendikten sonra hücreli ve hücresiz örnekler 27 C de 48 saat inkübe edildi. MTT solüsyonunun hazırlanması bölümünde açıklandığı gibi hazırlanan 10 mg/ml MTT solüsyonundan her kuyucuğa 10 µl eklendi ve 27 C de 4 saat inkübe edildi. Formazan kristalleri oluşumu ve hareketlerinin aktifliği invert mikroskopla tek tek incelendi ve fotoğrafları çekildi. MTT durdurucu solüsyondan 100 µl eklendi, formazan kristallerinin tamamen çözüldüğü mikroskobik olarak incelendi ve 30 dakika oda sıcaklığında beklenip ELIZA cihazı ile 570 nm dalga boyunda optik yoğunluk ölçüldü. 5.4 J774 Makrofaj Hücrelerinin Annexin-V&7-AAD İşaretlenmesi Apoptozis sırasında, hücre zarının fosfolipid iç tabakasında bulunan fosfatidilserinin (PS), hücre zarının dış tabakasına geçer. Bu yüzen Annexin V fosfatidilserine yüksek affinite ile bağlanarak apoptotik hücrelerin belirlenmesini sağlar. 7 -Amino-Aktinomisin D (7-AAD) ise,
56 42 DNA'nın sitozin ve guanin bazları arasına giren bir nükleik asit boyası olup canlı olan hücreleri canlı olmayanlardan ayırt edilmesinde kullanılır. J774 makrofaj kültürün için 6 kuyucuklu kültür plağı (TPP) kullanıldı. Thoma lamında hücrelerin sayımı yapıldıktan sonra 25x10 4 hücre olacak şekilde %10 FCS içeren RPMI 1640 medyumlu hücre süspansiyonundan eklendi. Makrofajların adhezyonu için 24 saat 37 C de %5 CO 2 içeren etüvde inkübasyonu sağlandı. 24 saat 37 C de %5 CO 2 içeren etüvde inkübasyona bırakılan 6 kuyucuklu kültür plağındaki makrofajlar üzerine, kontrol grubu ile birlikte 2,5 µg/ml, 250 µg/ml ve 2500 µg/ml olacak şekilde molekül ağırlığı 30,000 ve 100,000 olan PAA eklendi. Hücreler 37 C de %5 CO 2 içeren etüvde 48 saat inkübasyona bırakıldı. Hücreler flow sitometrik inceleme için Tripsin-EDTA ile plak üzerinden kaldırılıp 2 kez PBS ile yıkandı. Daha sonra hücrelere Annexin V-FITC çalışma solusyonundan 10 µl, 7- AAD çalışma solusyonundanda 5 µl ve bağlama tamponundanda 100 µl eklenerek 15 dakika oda sıcaklığında karanlıkta inkübasyona bırakıldı. 15 dakika inkübasyondan sonra her örneğe 900 µl daha bağlama tamponundan eklendi. Son olarak tüm örnekler 1 saat saat içinde flow sitometride analiz edildi. 5.5 Promastigotların Giemsa ile Boyanması MONI/EP126 promastigotları 5x10 5 parazit/ml olacak şekilde kültür tüplerine ekildi. 27 C soğutmalı etüvde 24 saat inkübasyona bırakıldıktan deney gruplarına 1, 5 ve 15 mg/ml olacak şekilde molekül ağırlığı 30,000 ve 100,000 olan PAA eklendi. Daha sonra PAA ya 1, 24 ve 48 saat maruz kalmış deney grupları ile kontrol gruplarından 100 µl alınarak alkolle (Riedel, 32221) temizlenmiş lam (Metzel gloser) üzerine yayma yapıldı. Lam üzerindeki örnek kurutuldu ve metanol (Merck,106009) ile 5 dakika oda sıcaklığında fikse edildi. Simir kurutuldu ve 1:10 oranında sulandırılmış Giemsa boyasında (Merck, ) 25 dakika oda sıcaklığında boyama işlemi gerçekleştirildi. Giemsa ile boyanmış örnekler çeşme suyu ile yıkandı, kurutuldu. Son olarak ışık mikroskobunda (100x) immersiyon yağı kullanılarak incelendi. 5.6 Makrofaj Hücrelerinin L. infantum ile Enfekte Edilmesi Makrofaj kültürünün enfekte edilmesi amacıyla L.infantum (MONI/EP126) kullanıldı. Makrofaj kültürünü promastigot ile enfekte etmek için 24 kuyucuklu kültür plağı (TPP) kullanıldı. Steril 12 mm çapındaki yuvarlak lameller tek tek kuyucuklara yerleştirildi. Thoma lamında hücrelerin sayımı yapıldı. Lamel yerleştirilen kuyucuklara 5x10 4 hücre olacak şekilde
57 43 RPMI 1640 (%10 FCS) içeren hücre süspansiyonundan eklendi. Makrofajların lamellerin üzerine adhezyonu için 24 saat 37 C de %5 CO 2 içeren etüvde inkübasyonu sağlandı. 24 saat 37 C de %5 CO 2 içeren etüvde inkübasyona bırakılan 24 kuyucuklu kültür plağındaki makrofajlar üzerine, molekül ağırlığı 30,000 ve 100,000 PAA ya 1, 24 ve 48 saat maruz kalmış deney grupları ile kontrol gruplarından hücre başına 10 promastigot düşecek şekilde (5x10 5 parazit) promastigotlar inoküle edildi. Hazırladığımız plate %5 CO 2 içeren etüvde 37 C de 4 saat süreyle inkübe edilerek makrofajların enfekte olması sağlandı. Daha sonra fagosite edilmeyen ekstrasellüler promastigotları uzaklaştırmak için 1 ml RPMI 1640 ile kuyular 3 er kez yıkandı ve yeni hazırlanmış RPMI 1640 (%10 FCS) besiyerinden 1 ml ilave edildi. Deney ve kontrol grubunda bulunan lameller 24. ve 48. saatlerde Giemsa boyanarak ışık mikroskobunda (100x) enfektiflik hesaplandı Enfekte Olmuş Makrofajların Giemsa ile Boyanması Enfekte makrofajlar 24 ve 48 saat 37 C de %5 CO 2 içeren etüvde inkübasyona bırakıldıktan sonra, 24 kuyucuklu kültür plağındaki besiyeri döküldü. Her kuyu 500 µl PBS ile bir kez yıkandı. PBS boşaltıldıktan sonra her kuyu 500 µl PBS-metanol (1:1,5) solusyonu ile yıkandı. PBS-metanol solusyonu boşaltıldıktan sonra her kuyuya 250 µl metanol eklendi ve oda sıcaklığında 10 dakika beklendi. Metanol döküldükten sonra tekrar metanol ile yıkama yapılır. Sonra hücre yüzeyini örtecek miktarda (200 µl) Giemsa eklendi. 24 lük kültür plağı hafifçe sallanarak boyanın tüm yüzeye yayılması sağlandı. 3 dakika oda sıcaklında duran boya döküldü. Boya döküldükten sonra 24 lük kültür plağı çeşme suyu altında yıkandı. Daha önceden 24 lük kültür plağı içine koyduğumuz 12 mm lik lameller çıkarılarak, Giemsa ile boyanmış olan enfekte makrofajlar ışık mikroskobunda x100 büyütmede immersiyon yağı (Applichem, 3w07849) ile gözlendi Enfektifliğin Hesaplanması Giemsa boyama ile elde edilen simirlerde toplam 200 makrofaj içinden kaçtanesinin enfekte olduğu belirlendi (yüzde enfektiflik). Daha sonra toplam amastogot sayısı belirlendi. Böylece ortalama bir makrofaja düşen amastigot sayısı hesplandı. Son olarak enfeksiyon indeksini (infection index) belirlemek için yüzde enfektiflik sayısı ile ortalama aamastigot sayısı çarpıldı.
58 İstatistik Analiz Deneyler en az 3 kez tekrarlandı. Sonuçlar ortalama ± SD olarak ifade edildi. Varyansların homojenitesi Levene's test ile belirlendi. Sonuçların değerlendirilmesinde parametrik testler (t student, varyans analizi, F, ve Tukey s post-hoc test) kullanıldı. Tüm veriler Statistical Packages of Social Sciences (SPSS, version 16.0 for Windows) programı ile yapıldı ve p<0,05 değeri istatistiki olarak anlamlı kabul edildi.

59 45 6. DENEYSEL SONUÇLAR VE TARTIŞMALAR 6.1 Poliakrilik Asitin J774 Hücrelerine Apoptotik Etkisinin İncelenmesi Molekül ağırlığı ve olan poliakrilik asitin J774 hücrelerine etkisi, Annexin- V&7-AAD işaretlenerek, flow sitometri ile incelendi. PAA bulunmayan kontrol grubunda 48 saat sonra hücre canlılığı % 94,47 (%5,53 nekrotik) olup, apoptotik hücre ölümü gözlenmemiştir (Şekil 6.1). 2,5 µg/ml ve molekül ağırlığında poliakrilik asit ile 48 saat inkübe edilen J774 hücrelerinin canlılığı PAA da %96,83 (%3,17 nekrotik) (Şekil 6.2), PAA da ise %97,55 (%2,45 nekrotik) (Şekil 6.3) olup, kontrol ile aralarında istatiski olarak fark görülmemiştir (p> 0,05). 250 µg/ml molekül ağırlığında poliakrilik asit ile 48 saat inkübe edilen J774 hücrelerinin canlılığı %84,12 (%15,88 nekrotik) olup apoptotik etki görülmemiştir (Şekil 6.4). 250 µg/ml molekül ağırlığında poliakrilik asit ile 48 saat inkübe edilen J774 hücrelerinin canlılığı %81,18 (%18,82 nekrotik) olup apoptotik etki görülmemiştir (Şekil 6.5) µg/ml molekül ağırlığında poliakrilik asit ile 48 saat inkübe edilen J774 hücrelerin %99,99 u nekrotik olarak ölmesine rağmen apoptotik etki görülmemiştir (p<0,05) (Şekil 6.6) µg/ml molekül ağırlığında poliakrilik asit ile 48 saat inkübe edilen J774 hücrelerinin %99,99 u nekrotik olarak ölmesine rağmen apoptotik etki tespit edilmemiştir (p<0,05) (Şekil 6.7). Şekil 6.1 PAA bulunmayan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası)
60 46 Şekil 6.2 PAA (2,5 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) 2,5 Şekil 6.3 PAA (2,5 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası)
61 47 Şekil 6.4 PAA (250 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) Şekil 6.5 PAA (250 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası)
 bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) 6.")
62 48 Şekil 6.6 PAA (2500 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) Şekil 6.7 PAA (2500 µg/ml MA: ) bulunan kültür ortamında J774 hücrelerinin Apoptotik-Nekrotik etkisinin flow sitometrik tayini (48 saat inkübasyon sonrası) 6.2 PAA nın J774 Hücre Kültüründe Toksik Etkisinin İncelenmesi Farklı molekül ağırlıklı polimerlerin toksik etkisini incelemek amacıyla, 96 kuyucuklu plağın her bir kuyucuğuna 10,000 J774 hücresi ekilip 24 saat inkübe edildi. Daha sonra her bir kuyucuğa MTT boyası ilave edildi ve hücre canlılığını belirlemek için formazan kristallerinin oluşumu ters mikroskopta incelendi. MTT bulunmayan (Şekil 6.8) ve MTT bulunan (Şekil 6.9) kültür ortamında J774 hücrelerinin morfolojileri gösterilmiştir. Şekil 6.8 de görüldüğü gibi hücreler kültür alanını kaplamış, morfolojik olarak intakt ve normaldir.



 ve 48 saatlik")
63 49 Şekil 6.8 J774 hücrelerinin 48 saat inkübasyon sonrası morfolojik görüntüsü (MTT eklenmeden önce) (20x). Şekil 6.9 J774 hücrelerinde formazan kristallerinin oluşumu (48 saat inkübasyon sonrası, MTT eklendikten sonra) (20x). MTT bulunan kültür ortamında ise formazan kristallerinin oluşumu net bir şekilde görülmektedir. Elde edilen sonuçlarda, kullanılan hücre miktarı (10,000 hücre/kuyucuk) ve 48 saatlik inkübasyon süresi ile hücrelerin kuyucuklarda normal bir şekilde adezyon yapıp çoğaldığı ve canlılıklarının yüksek olduğu görüldü. Bu nedenle farklı molekül ağırlığındaki polimerlerin kültür ortamındaki toksik etkisini incelemek için çalışmalarımızda bu prosedür uygulandı.


64 Molekül Ağırlığı Olan Poliakrilik Asitin J774 Hücreleri Üzerindeki Toksik Etkisinin İncelenmesi Poliakrilik asitin farklı konsantrasyonları (2,5 µg/ml, 5 µg/ml, 10 µg/ml, 25 µg/ml, 50 µg/ml, 100 µg/ml, 250 µg/ml, 500 µg/ml, 1000 µg/ml 2500 µg/ml) J774 hücre kültürlerinde 48 saat inkübe edildikten sonra PAA nın toksik etkisi MTT yöntemi ile morfolojik ve sayısal olarak incelendi. Şekil 6.10 da PAA nın en yüksek konsantrasyonu bulunan kültür ortamında hücrelerin mikroskobik görüntüleri verilmiştir. Hücreler atipik forma dönüşmüş, büzülüp küçülmüş, yüzey uzantılarını kaybetmiş ve hücreler arası alanda boşluklar oluşmuştur. Şekil 6.10 PAA nın 2500 µg/ml konsantrasyonunun J774 hücreleri üzerindeki sitopatik etkisi (20x). PAA (2500 µg/ml) bulunan kültür ortamında oluşan morfolojik değişiklikler 500 µg/ml ve 250 µg/ml lik konsantrasyonlarda da tespit edilmiştir (Şekil 6.11, 6.12).




65 51 Şekil 6.11 PAA nın 500 µg/ml konsantrasyonunun J774 hücreleri üzerindeki sitopatik etkisi (20x). Şekil 6.12 PAA nın 250 µg/ml konsantrasyonunun J774 hücreleri üzerindeki sitopatik etkisi (20x). Ancak en düşük konsantrasyon olan 2,5 µg/ml de diğer konsantrasyonlardan farklı olarak formazan kristali oluşmuş, ancak morfolojik olarak kontrole göre önemli bir değişiklik göstermemiştir (Şekil 6.13).

. Çizelge 6.")
66 52 Şekil 6.13 PAA nın 2,5 µg/ml konsantrasyonunun J774 hücreleri üzerindeki sitopatik etkisi (20x). Çizelge 6.1 de PAA nın J774 hücreleri üzerindeki toksik etkisi konsantrasyona bağlı olarak verilmiştir. Çizelgede görüldüğü gibi PAA nın konsantrasyonu arttıkça J774 hücrelerinin canlılıkları azalmıştır. En yüksek konsantrasyon olan 2500 µg/ml lik PAA bulunan ortamdaki hücre canlığı, kontrole göre anlamlı bir düşüş göstermiştir (P<0,05). Çizelge 6.1 PAA nın (MA:100,000 ) farklı konsantrasyonlarının J774 hücrelerinin canlılığına etkisi J774 0,6 Optik Yoğunluk 0,5 0,4 0,3 0,2 0,1 0 kontrol 2, Konsantrasyon (µg/ml)


67 Molekül Ağırlığı 30,000 Olan Poliakrilik Asitin J774 Hücreleri Üzerindeki Toksik Etkisinin İncelenmesi J774 hücreleri PAA nın farklı konsantrasyonları (2,5 µg/ml, 5 µg/ml, 10 µg/ml, 25 µg/ml, 50 µg/ml, 100 µg/ml, 250 µg/ml, 500 µg/ml, 1000 µg/ml, 2500 µg/ml) ile 48 saat inkübe edildi. PAA nın incelenen konsantrasyonlarının hücreler üzerindeki toksik etkisi mikroskobik ve MTT yöntemi ile sayısal olarak incelendi. Şekil 6.14 te PAA nın 2500 µg/ml konsantrasyonunun J774 hücrelerinde oluşturduğu morfolojik değişiklikler verilmiştir. Hücrelerde yuvarlaklaşma, küçülme, uzantılarını kaybetme gibi sitopatik değişiklikler meydana gelmiştir. Ayrıca hücrelerde formazan kristali oluşumu da olduğu görülmüştür. Şekil 6.14 PAA nın 2500 µg/ml konsantrasyonun J774 hücreleri üzerindeki sitopatik etkisi (20x).




68 54 Şekil 6.15 PAA nın 1000 µg/ml konsantrasyonun J774 hücreleri üzerindeki sitopatik etkisi (20x). Şekil 6.16 da PAA nın 250 µg/ml lik konsantrasyonu bulunan kültür ortamında hücrelerde meydana gelen morfolojik değişiklikler gösterilmiştir. PAA nın 1000 ve 2500 µg/ml lık konsantrasyondan farklı olarak 250 µg/ml de hücrelerin tamamının canlılığını kaybetmediği görülmektedir. Şekil 6.16 PAA nın 250 µg/ml konsantrasyonun J744 hücreleri üzerindeki sitopatik etkisi (20x). PAA nın en düşük konsantrasyonunda (2,5 µg/ml) ise hücrelerde kontrole kıyasla morfolojik olarak değişiklikler görülmemiştir (Şekil 6.17).
 toksik etkinin olmadığı ve hücrelerin canlılığının azalmadığı görülmektedir.")
. Çizelge 6.")
69 55 Şekil 6.17 PAA nın 2,5 µg/ml konsantrasyonun J774 hücreleri üzerindeki sitopatik etkisi (20x). Çizelge 6.2 de ise PAA nın J774 hücreleri üzerindeki etkisi kantitatif olarak verilmiştir. Çizelgeden görüldüğü gibi, en düşük konsantrasyonda (2,5 µg/ml) toksik etkinin olmadığı ve hücrelerin canlılığının azalmadığı görülmektedir. Buna karşın, 250 µg/ml den itibaren artan konsantrasyonlar ile hücrelerin canlılıkları azalmaya başlamıştır. En yüksek konsantrasyonda (2500 µg/ml) canlılık tamamen ortadan kalkmıştır. En yüksek konsantrasyonda hücrelerin kontrole göre canlılıkları anlamlı ölçüde düşmüştür (p<0,05). Çizelge 6.2 PAA nın (MA:30,000) farklı konsantrasyonlarının J774 hücrelerinin canlılığına etkisi J774 0,7 Optik Yoğunluk 0,6 0,5 0,4 0,3 0,2 0,1 0 kontrol 2, Konsantrasyon (µg/ml)

70 56 Sonuçlar PAA nın 100 µg/ml den yüksek olan konsantrasyonlarda, J774 hücreleri üzerinde toksik etkisi olduğunu göstermektedir. 6.3 PAA nın Leishmania promastigotlarına Toksik Etkisinin İncelenmesi Leishmania promastigotları üzerine poliakrilik asitin toksik etkisini MTT yöntemiyle incelemek amacı ile 96 kuyucuklu plağın her bir kuyusuna 30,000 promastigot ekilip, 48 saat inkübe edildi. Ayrıca Giemsa boyama yöntemiyle promastigotlardan alınan örneklerin morfolojik yapıları incelendi. Promastigotların hareketliliği ise invert mikroskop altında incelendi. Yapılan incelemelerde promastigotların morfolojik yapıları normal olup, kinetoplast, çekirdek ve kamçılarının oldukça belirgin olduğu gözlendi. (Şekil 6.18). Şekil 6.18 L.infantum promastigotlarının morfolojik görüntüsü (Giemsa boyama, 100x) (48 saat inkübasyon sonrası) Molekül Ağırlığı 100,000 Olan Poliakrilik Asitin Ep126 Promastigotları Üzerindeki Toksik Etkisinin İncelenmesi 96 kuyucuklu plağın her bir kuyucuğuna 30,000 promastigot ekilip, 48 saat inkübe edildikten sonra poliakrilik asitin (MA: 100,000 ) farklı konsantrasyonları (1 mg/ml, 5 mg/ml ve 15 mg/ml) Ep126 promastigotları üzerine eklenerek 48 saat inkübasyona bırakıldı. Promastigotlara verilen en yüksek konsantrasyon olan 15 mg/ml PAA bulunan kültür ortamında, polimer eklenmesinden hemen sonra hücrelerin kontrole göre hareketliliklerinde önemli ölçüde azalma tespit edildi. Morfolojik inceleme amacıyla 100,00 PAA ya 1 saat, 24

 ayırt edilemediği gözlendi (Şekil 6.19).")
.")
. Şekil 6.")
71 57 saat ve 48 saat maruz bırakılan promastigotlardan alınan örneklerden Giemsa boyama yapılarak promastigotların morfolojik yapıları incelendi. 15 mg/ml PAA ya 1 saat maruz kalmış promastigotların hücre bütünlüğünü kaybettiği ve hücre içi yapılarının (kinetoplast, çekirdek) ayırt edilemediği gözlendi (Şekil 6.19). 15 mg/ml PAA ya 24 saat maruz kalmış promastigotların da hücre bütünlüğünü kaybettiği fakat hücre içi yapıların tam olarak olmasa da görüldüğü tespit edildi (Şekil 6.20). 15 mg/ml PAA ya 48 saat maruz kalmış promastigotlarda ise, hücre bütünlüğü tamamen kaybolmuş fakat kültürde yapılan incelemelerde, halen canlılığını koruyan promastigotların olduğu gözlendi (Şekil 6.21). Şekil mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) Şekil mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x)
 5 mg/ml PAA ya 1 saat maruz kalmış promastigotlar kontrole göre atipik olup, çekirdekleri")

72 58 Şekil mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) 5 mg/ml PAA ya 1 saat maruz kalmış promastigotlar kontrole göre atipik olup, çekirdekleri görülmesine rağmen kinetoplast görülmemektedir (Şekil 6.22). 5 mg/ml PAA ya 24 saat maruz kalmış promastigotların kontrole göre atipik olmasına rağmen, polimere bir saat maruz kalmış hücrelere göre daha intakt oldukları gözlendi (Şekil 6.23). 5 mg/ml PAA ya 48 saat maruz kalmış promastigotlarda ise, parazitler atipik olup kamçıları görülmemektedir. Ancak parazitlerin 24. saate göre daha iyi durumda olduğu gözlendi (Şekil 6.24). Şekil mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x)
 1 mg/ml PAA ya 1")
.")
.")
73 59 Şekil mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) Şekil mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) 1 mg/ml PAA ya 1 saat maruz kalmış promastigotların hücre bütünlüğü kaybolmuş olup, hücre içi yapıların ayırt edilemediği gözlendi (Şekil 6.25). 1 mg/ml PAA ya 24 saat maruz kalmış promastigotların durumları, birinci saate göre daha iyi olup, hücre içi yapılar ayırt edilebilmektedir (Şekil 6.26). 1 mg/ml PAA ya 48 saat maruz kalmış promastigotların morfolojilerinin ise, 24. saatteki morfolojiye benzer olduğu gözlendi (Şekil 6.26).


74 60 Şekil mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) Şekil mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x)

.")
75 61 Şekil mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) MTT yöntemi sonucu elde edilen formazan kristalleri çözündürülerek optik yoğunluğu ölçülmüştür. Kontrolün canlılığı %100 kabul edilerek Çizelge 6.3 te PAA nın farklı konsantrasyonlarının Ep126 promastigotları üzerine etkisi verilmiştir. 1 mg/ml PAA ya maruz kalmış promastigotların canlılığı ile kontrol arasında istatiski olarak fark yoktur (p> 0,05). 5 mg/ml PAA ya maruz kalmış promastigotların kontrole göre canlılığı anlamlı ölçüde yarı yarıya düşmüştür (p<0,05). 5 mg/ml nin üzerinde PAA ya maruz kalmış promastigotlarda canlılık giderek azalmıştır. Fakat kültür ortamında polimerin etkisi incelendiğinde, promastigotların 15 mg/ml gibi yüksek konsantrasyonlarda da canlılığını koruduğu ve 48. saatten sonra yapılan sayımlarda kontrole göre hücre sayısında artış başladığı görülmüştür (Çizelge 6.4). Elde edilen MTT sonuçlarına göre 100,000 PAA da IC 50 4,79 mg/ml dir. Çizelge 6.3 MA:100,000 olan PAA nın Ep126 promastigotlarının canlılığına etkisi % Canlılık Konsantrasyon (mg/ml)
76 62 Çizelge 6.4 MA:100,000 olan PAA nın Ep126 promastigotlarına in vitro etkisi Parazit Sayısı (/ml) Zaman (saat) Kontrol 1 mg/ml 5 mg/ml 15 mg/ml Molekül Ağırlığı 30,000 Olan Poliakrilik Asitin Ep126 Promastigotları Üzerindeki Toksik Etkisinin İncelenmesi 96 kuyucuklu plağın her bir kuyucuğuna 30,000 promastigot ekilip, 48 saat inkübe edildikten sonra, poliakrilik asitin (MA: 30,000 ) farklı konsantrasyonları (0,1 mg/ml, 1 mg/ml, 5 mg/ml ve 15 mg/ml) Ep126 promastigotları üzerine eklenerek 48 saat inkübasyona bırakıldı. Promastigotlara verilen en yüksek konsantrasyon olan 15 mg/ml PAA bulunan kültür ortamında, polimer eklenmesinden hemen sonra hücrelerin kontrole göre hareketliliklerinde önemli ölçüde azalma tespit edildi. Morfolojik inceleme amacıyla 30,000 PAA ya 1 saat, 24 saat ve 48 saat maruz bırakılan promastigotlardan alınan örneklerden Giemsa boyama yapılarak, promastigotların morfolojik yapıları incelendi. 15 mg/ml PAA ya 1 saat maruz kalmış promastigotların hücre bütünlüğünü kaybettiği ve hücre içi yapılarının (kinetoplast, çekirdek) ayırt edilemediği gözlendi (Şekil 6.28). 15 mg/ml PAA ya 24 saat maruz kalmış promastigotların da hücre bütünlüğünü kaybettiği, fakat promastigot çekirdeği ile kamçılarının belirgin olduğu gözlendi (Şekil 6.29). 15 mg/ml PAA ya 48 saat maruz kalmış promastigotlarda ise hücre bütünlüğünün tamamen kaybolduğu fakat kültürden yapılan incelemelerde canlılığını koruyan promastigotların olduğu gözlendi (Şekil 6.30). Elde edilen MTT sonuçlarına göre30,000 PAA da IC 50 23,78 mg/ml dir.



77 63 Şekil mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) Şekil mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x)
 5")
.")


78 64 Şekil mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) 5 mg/ml PAA ya 1 saat maruz kalmış promastigotlar kontrole göre atipik olup çekirdek, kinetoplast ve kamçıları görülebilmektedir (Şekil 6.31). 5 mg/ml PAA ya 24 saat maruz kalmış promastigotlar kontrole göre atipik olup hücre bütünlüğünü tamamen kaybetmeye başlamıştır. Hücre içi yapıların belirgin olmadığı görüldü (Şekil 6.32). 5 mg/ml PAA ya 48 saat maruz kalmış promastigotlar ise atipik olup parazitlerin 24. saate göre daha iyi durumda olduğu ve bazı parazitlerin bölünebildiği gözlendi (Şekil 6.33). Şekil mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x)
 1 mg/ml PAA ya 1 saat maruz")
.")
.")
79 65 Şekil mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) Şekil mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları(giemsa boyama, 100x) 1 mg/ml PAA ya 1 saat maruz kalmış promastigotların hücre bütünlüğü tamamen kaybolmuş olup, hücre içi yapıların ayırt edilemediği gözlendi (Şekil 6.34). 1 mg/ml PAA ya 24 saat maruz kalmış promastigotların durumları birinci saate göre daha iyi olup, hücre içi yapılar ayırt edilebilmektedir (Şekil 6.35). 1 mg/ml PAA ya 48 saat maruz kalmış promastigotların neredeyse kontrol ile aynı morfolojiye sahip olduğu, kinetoplast, kamçı ve çekirdeklerinin ayırt edilebildiği görülmektedir (Şekil 6.36).

 Şekil 6.")
")
80 66 Şekil mg/ml PAA ya 1 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) Şekil mg/ml PAA ya 24 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x)
.")
81 67 Şekil mg/ml PAA ya 48 saat maruz kalmış Ep126 promastigotları (Giemsa boyama, 100x) 1 mg/ml, 5 mg/ml ve 15 mg/ml PAA ya maruz kalmış promastigotların canlılığı 48. saatte kontrole göre anlamlı ölçüde düşmüştür (p<0,05). Fakat kültür ortamında polimerin etkisi incelendiğinde, promastigotların, 15 mg/ml gibi yüksek konsantrasyonlarda da canlılığını koruduğu ayrıca 48. saatten sonra yapılan sayımlarda kontrole göre hücre sayısında artış başladığı görüldü (Çizelge 6.5). Çizelge 6.5 MA:30,000 olan PAA nın Ep126 promastigotlarına in vitro etkisi Parazit Sayısı (/ml) Zaman (saat) Kontrol 1 mg/ml 5 mg/ml 15 mg/ml
82 Poliakrilik Asite Maruz Kalmış Ep126 (L. Infantum) Promastigotlarının J774 Makrofaj Hücreleri ile Etkileşimi ve Enfektifliğinin İncelenmesi Parazitler 1 mg/ml, 5 mg/ml ve 15 mg/ml PAA ya 1saat, 24 saat ve 48 saat maruz bırakıldıktan sonra 2 kez PBS ile yıkanıp sayım yapılarak, J774 makrofaj hücrelerine, makrofaj başına 10 promastigot olacak şekilde verildikten 4 saat sonra, makrofajlara giremeyen promastigotlar, yıkama yapılarak ortamdan uzaklaştırıldı. Daha sonra enfektifliği belirlemek amacıyla 24. ve 48. saatlerde Giemsa boyama yapıldı. Genel olarak elde edilen sonuçlarda, polimere maruz kalmış ortamlarda kontrole göre enfektifliğin önemli ölçüde düştüğü gözlendi. Giemsa boyama ile elde edilen simirlerde toplam 200 makrofaj içinden kaç tanesinin enfekte olduğu belirlendi (yüzde enfektiflik). Daha sonra toplam amastigot sayısı belirlendi. Böylece ortalama bir makrofaja düşen amastigot sayısı hesplandı. Son olarak enfeksiyon indeksini (infection index) belirlemek için yüzde enfektiflik sayısı ile ortalama amastigot sayısının çarpımı hesaplandı. Bu hesaplamalara göre, polimere maruz kalmamış 24. saatteki kontrol grubunda makrofajların %70,27 si enfekte makrofaj olup, ortalama bir makrofaja düşen promastigot sayısı ise 3.1 dir. Bu durumda infeksiyon indeksi 70,27 x 3,1 = 218 dir. Polimere maruz kalmamış 48. saatteki kontrol grubunda makrofajların %28,12 si enfekte makrofaj olup, ortalama bir makrofaja düşen promastigot sayısı ise 0,6 dır. Bu durumda infeksiyon indeksi 28,12 x 0,6 = 19 dur. 48. saatte enfektifliğin bu kadar düşmesinin nedeni; in vitro koşullarda makrofaj sayısının sabit olması ve amastigotların diğer makrofajları enfekte etmesi için yeterli zamanı in vitro da sağlayamaması olabilir. Şekil 6.37 de Giemsa boyama ile kontrol grubundaki enfekte makrofajlar görülmektedir.
 promastigotları molekül ağırlığı 100,000 1 mg/ml, 5 mg/ml ve 15 mg/ml PAA ya 1 saat, 24 saat ve 48 saat maruz bırakıldıktan sonra J774 makrofaj hücrelerine olan etkisi incelendi. 24. ve 48. saatlerde Giemsa boyama yapılarak enfektiflik hesaplandı.")

83 69 Şekil 6.37 Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) Poliakrilik Asite (MA:100,000) Maruz Kalmış Ep126 Promastigotlarının J774 Hücreleri ile Etkileşimi ve Enfektifliğinin İncelenmesi Ep126 (L. Infantum) promastigotları molekül ağırlığı 100,000 1 mg/ml, 5 mg/ml ve 15 mg/ml PAA ya 1 saat, 24 saat ve 48 saat maruz bırakıldıktan sonra J774 makrofaj hücrelerine olan etkisi incelendi. 24. ve 48. saatlerde Giemsa boyama yapılarak enfektiflik hesaplandı PAA ya (15 mg/ml, MA:100,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi 15 mg/ml konsantrasyona 1saat, 24 saat ve 48 saat maruz kalmış tüm deney gruplarında 24. saat Giemsa örneklerinde enfekte olmuş makrofajlardaki amastigotlar oldukça küçük ve kontrole göre atipiktir. 15 mg/ml PAA'ya 24 ve 48 saat maruz kalmış gruplarda 24. saat Giemsa örneklerinde enfekte makrofaj görülmezken (Şekil 38 ve Şekil 39), 1 saat maruz kalmış gruplarda enfektiflik %1 in altındadır (Şekil 40).



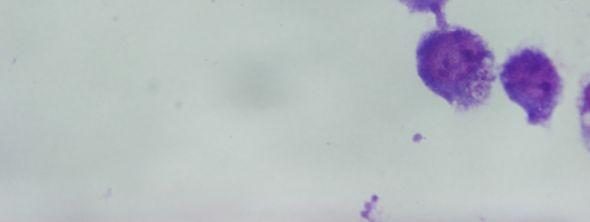
84 70 Şekil mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) Şekil mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x)

 15 mg/ml polimere maruz kalmış tüm gruplarda, 48.")

.")
85 71 Şekil mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) 15 mg/ml polimere maruz kalmış tüm gruplarda, 48. saatte Giemsa örneklerinde enfektiflik %1 in altında olup, makrofajların kültür ortamındaki 48. saat morfolojilerinin normal ve canlılıklarının 24. saate göre daha iyi olduğu gözlendi (Şekil 41, Şekil 42 ve Şekil 43). Şekil mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x)


 6.4.1.")

86 72 Şekil mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) Şekil mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) PAA ya (5 mg/ml, MA:100,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi 5 mg/ml konsantrasyona 1saat, 24 saat ve 48 saat maruz kalmış tüm deney gruplarında 24. saat Giemsa örneklerinde enfekte olmuş makrofajlardaki amastigotlar hücre bütünlüğünü kaybetmiş, oldukça küçük olup, amastigot çekirdeği ve kinetoplastı belirgin değildir. 15 mg/ml polimere 24 ve 48 saat maruz kalmış gruplarda 24. saat Giemsa örneklerinde enfekte
.")
. Şekil 6.")


87 73 makrofaj %1 in altında olup, amastigotlarda kontrole göre atipiktir (Şekil 44 ve Şekil 45). Polimere 1 saat maruz kalmış gruplarda ise enfekte makrofaj görülmemiştir (Şekil 46). Şekil mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) Şekil mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x)

 5 mg/ml polimere maruz kalmış tüm gruplarda, 48.")


88 74 Şekil mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) 5 mg/ml polimere maruz kalmış tüm gruplarda, 48. saat Giemsa örneklerinde enfekte makrofaj sayısı kontrole göre çok az olup, enfekte makrofajlardaki amastigotların tamamı hücre bütünlüğünü kaybetmiştir (Şekil 47, Şekil 48 ve Şekil 49). Şekil mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x)


 6.4.1.")

89 75 Şekil mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) Şekil mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) PAA ya (1 mg/ml, MA:100,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi 1 mg/ml konsantrasyona 48 saat maruz kalmış deney grubunda, 24. saat Giemsa örneklerinde enfektiflik %46 ve makrofaj başına düşen amastigot sayısıda 2,2 dir. Enfekte makrofajlarda bulunan amastigotların bir kısmı morfolojik olarak kontrole yakın olurken, bazı amastigotlar ise hücre bütünlüğünü kaybetmiş, oldukça küçük olup çekirdek ve kinetoplastı belirgin değildir (Şekil 50). 1 mg/ml polimere 1 ve 24 saat maruz kalmış gruplarda 24. saat Giemsa

. Şekil 6.")


90 76 örneklerinde enfektiflik yaklaşık %18 ve makrofaj başına düşen amastigot sayısıda 0,4 tür. Amastigotlarda kontrole göre atipiktir (Şekil 51 ve Şekil 52). Şekil mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) Şekil mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x)




91 77 Şekil mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) 1 mg/ml polimere maruz kalmış tüm gruplarda, 48. saat Giemsa örneklerinde enfektiflik kontrolün 24. ve 48. saatleri arasındaki farkla kıyaslandığında önemli ölçüde düşmüştür. Ayrıca kontrolde 48. saatte amastigotlar normal morfoloji gösterirken, 1 mg/ml polimere maruz kalmış tüm gruplarda amastigotların tamamı hücre bütünlüğünü kaybetmiştir (Şekil 53, Şekil 54 ve Şekil 55). Şekil mg/ml 100,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x)



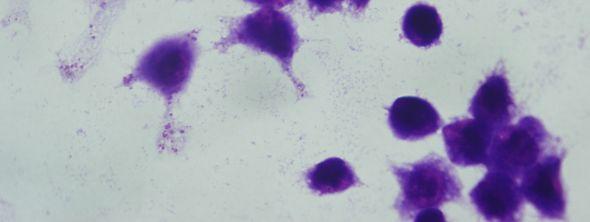
92 78 Şekil mg/ml 100,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) Şekil mg/ml 100,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x)
93 79 Çizelge 6.6 Poliakrilik Asite (MA:100,000) Maruz Kalmış Ep126 Promastigotlarının J774 Hücrelerine Enfektifliği MA: 100,000 PAA 24 Saat Sonra Enfektiflik 48 Saat Sonra Enfektiflik % Enfektiflik Amastigot Sayısı Enfeksiyon İndeksi % Enfektiflik Amastigot Sayısı Enfeksiyon İndeksi Kontrol 70,27 3, ,12 0, mg/ml, 1 Saat 17,2 0, , mg/ml, 24 Saat 17,7 0, , mg/ml, 101,2 48 Saat 46 2,2 (amastigotlar 2 0,01 0 atipik) 5 mg/ml, 1 Saat 1 0, , mg/ml, 24 Saat 4 0, , mg/ml, 48 Saat 15 mg/ml, 1 Saat 15 mg/ml, 24 Saat 15 mg/ml, 48 Saat 1 0, , , , , , , , Poliakrilik Asite (MA:30,000) Maruz Kalmış Ep126 Promastigotlarının J774 Hücreleri ile Etkileşimi ve Enfektifliğinin İncelenmesi Ep126 (L. Infantum) promastigotları molekül ağırlığı 30,000 olan 1 mg/ml, 5 mg/ml ve 15 mg/ml PAA ya 1saat, 24 saat ve 48 saat maruz bırakıldıktan sonra J774 makrofaj hücrelerine olan etkisi incelendi. 24. ve 48. saatlerde Giemsa boyama yapılarak enfektiflik hesaplandı PAA ya (15 mg/ml, MA:30,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi 15 mg/ml konsantrasyona 1 saat maruz kalmış deney grubunda, 24. saat Giemsa örneklerinde enfektiflik %21 ve makrofaj başına düşen amastigot sayısıda 1,5 tir. Enfekte makrofajlarda bulunan amastigotların morfolojik olarak kontrole yakın olsada çekirdek ve kinetoplastları tam olarak belirgin değildir (Şekil 6.56). Polimere 48 ve 24 saat maruz kalmış gruplarda, 24.
 çok az olup, amastigotlarda kontrole göre atipiktir")

 Şekil 6.")
")
94 80 saat Giemsa örneklerinde enfekte makrofaj sayısı (< %1) çok az olup, amastigotlarda kontrole göre atipiktir (Şekil 57 ve Şekil 58). Şekil mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) Şekil mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x)




95 81 Şekil mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) 15 mg/ml polimere 1saat maruz kalmış, 48. saat Giemsa örneklerinde, enfektiflik kontrolün 24. ve 48. saatleri arasındaki farkla kıyaslandığında önemli ölçüde düşmüştür. Ayrıca kontrolde 48. saatte amastigotlar normal morfoloji gösterirken, 15 mg/ml polimere maruz kalmış tüm gruplarda amastigotların tamamı hücre bütünlüğünü kaybetmiştir (Şekil 59). Polimere 24 ve 48 saat maruz kalmış gruplarda, 48. saat Giemsa örneklerinde enfekte makrofaj sayısı (< %1) çok az olup, amastigotlarda kontrole göre atipiktir (Şekil 60 ve Şekil 61). Şekil mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x)


 Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi 5")
 çok az olup, amastigotlarda kontrole göre")
96 82 Şekil mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) Şekil mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) PAA ya (5 mg/ml, MA:30,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi 5 mg/ml konsantrasyona 24 ve 48 saat maruz kalmış deney grubunda, 24. saat Giemsa örneklerinde enfekte makrofaj sayısı (< %1) çok az olup, amastigotlarda kontrole göre atipiktir (Şekil 62 ve Şekil 63). 5 mg/ml konsantrasyona 1 saat maruz kalmış örneklerde enfekte makrofaj sayısı 24. ve 48. saate göre fazla olsada, amastigotlar atipik olup, çekirdek ve kinetoplastları tam olarak belirgin değildir (Şekil 6.64).



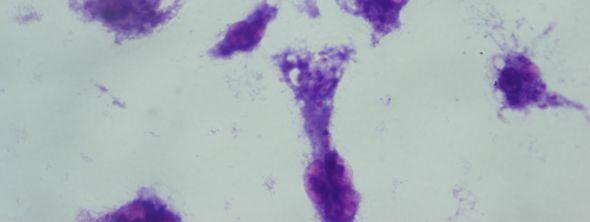
97 83 Şekil mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) Şekil mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x)


.")

98 84 Şekil mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) 5 mg/ml polimere maruz kalmış tüm deney gruplarında, 48. saat Giemsa örneklerinde, enfekte makrofaj sayısı (< %1) çok az olup, amastigotlarda kontrole göre atipiktir (Şekil 65, Şekil 66 ve Şekil 67). Ayrıca tüm deney gruplarında makrofaj morfolojisi normal iken, makrofaj canlılığı 24. saate göre artmıştır. Şekil mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x)


 Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile")

99 85 Şekil mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) Şekil mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) PAA ya (1 mg/ml, MA:30,000) Maruz Kalmış Promastigotların J774 Makrofaj Hücreleri ile Etkileşimi 1 mg/ml konsantrasyona maruz kalmış tüm deney grubuplarında, 24. saat Giemsa örneklerinde enfektiflik %3 olup, amastigotlarda kontrole göre atipiktir. (Şekil 67, Şekil 68 ve Şekil 69). Amastigot canlılığı özellikle polimere 1 saat maruz kalmış deney grubunda dikkat çekmektedir. Amastigotların çekirdek ve kinetoplastları tam olarak belirgin değildir.



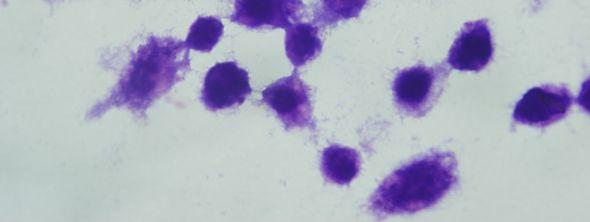
100 86 Şekil mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) Şekil mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x)




101 87 Şekil mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 24 saat sonra, 100x) 1 mg/ml konsantrasyona maruz kalmış tüm deney grubuplarında, 48. saat Giemsa örneklerinde enfektiflik 24. saate göre önemli ölçüde azalmıştır. Özellikle polimere 1 saat maruz tutulan deney grubunda enfekte makrofaj görülmemeiştir. Çok az sayıda enfekte makrofajlardaki amastigotlar tamamen deforme olmuştur (Şekil 70, Şekil 71 ve Şekil 72). Şekil mg/ml 30,000 PAA ya 48 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x)



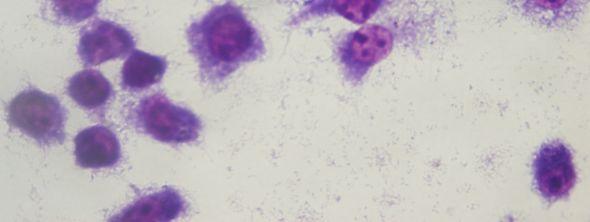
102 88 Şekil mg/ml 30,000 PAA ya 24 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x) Şekil mg/ml 30,000 PAA ya 1 saat maruz kalmış Ep126 promastigotları ile enfekte olmuş J774 makrofaj hücreleri (Giemsa boyama, 48 saat sonra, 100x)
Global Leishmaniasis. Leishmaniasis. Türkiye de leishmaniasis. Leishmaniasis. Leishmaniasis
 Leishmaniasis Leishmaniasis Leishmaniasis Tropik ve subtropik iklimlerde, Akdeniz bölgesi ülkelerde (88 ülke) görülür Her yıl 1,5 milyon yeni olgu eklenmektedir 350 milyon insan risk altında 23 farklı
Leishmaniasis Leishmaniasis Leishmaniasis Tropik ve subtropik iklimlerde, Akdeniz bölgesi ülkelerde (88 ülke) görülür Her yıl 1,5 milyon yeni olgu eklenmektedir 350 milyon insan risk altında 23 farklı
Hidrazon Yapısındaki On Adet Bileşiğin Antileishmanial Aktivitesinin Araştırılması
 Hidrazon Yapısındaki On Adet Bileşiğin Antileishmanial Aktivitesinin Araştırılması Şahin Direkel 1, Seda Tezcan 1, Semra Utku 2, Gönül Aslan 1, Mehtap Gökçe 3, M. Fethi Şahin 3, Nuran Delialioğlu 1, Mahmut
Hidrazon Yapısındaki On Adet Bileşiğin Antileishmanial Aktivitesinin Araştırılması Şahin Direkel 1, Seda Tezcan 1, Semra Utku 2, Gönül Aslan 1, Mehtap Gökçe 3, M. Fethi Şahin 3, Nuran Delialioğlu 1, Mahmut
Prof. Dr. Ahmet Özbilgin Celal Bayar Üniversitesi Tıp Fakültesi Parazitoloji Anabilim Dalı,Manisa
 Prof. Dr. Ahmet Özbilgin Celal Bayar Üniversitesi Tıp Fakültesi Parazitoloji Anabilim Dalı,Manisa 1. REDUCTION (Azaltma) 2. REPLACEMENT (Yerine bir şey koyma) 3. REFİNEMENT (Hayvanları rahatının sağlanması)
Prof. Dr. Ahmet Özbilgin Celal Bayar Üniversitesi Tıp Fakültesi Parazitoloji Anabilim Dalı,Manisa 1. REDUCTION (Azaltma) 2. REPLACEMENT (Yerine bir şey koyma) 3. REFİNEMENT (Hayvanları rahatının sağlanması)
ADIM ADIM YGS LYS Adım DOLAŞIM SİSTEMİ 5 İNSANDA BAĞIŞIKLIK VE VÜCUDUN SAVUNULMASI
 ADIM ADIM YGS LYS 177. Adım DOLAŞIM SİSTEMİ 5 İNSANDA BAĞIŞIKLIK VE VÜCUDUN SAVUNULMASI İNSANDA BAĞIŞIKLIK VE VÜCUDUN SAVUNULMASI Hastalık yapıcı organizmalara karşı vücudun gösterdiği dirence bağışıklık
ADIM ADIM YGS LYS 177. Adım DOLAŞIM SİSTEMİ 5 İNSANDA BAĞIŞIKLIK VE VÜCUDUN SAVUNULMASI İNSANDA BAĞIŞIKLIK VE VÜCUDUN SAVUNULMASI Hastalık yapıcı organizmalara karşı vücudun gösterdiği dirence bağışıklık
T.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
 T.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ İSTANBUL DAKİ KAN MERKEZLERİNDEN ELDE EDİLEN DONÖR KAN ÖRNEKLERİNDE, LEISHMANIA PARAZİTLERİNİN YENİ MİKROKÜLTÜR YÖNTEMİ VE DİĞER YÖNTEMLER İLE KARŞILAŞTIRMALI
T.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ İSTANBUL DAKİ KAN MERKEZLERİNDEN ELDE EDİLEN DONÖR KAN ÖRNEKLERİNDE, LEISHMANIA PARAZİTLERİNİN YENİ MİKROKÜLTÜR YÖNTEMİ VE DİĞER YÖNTEMLER İLE KARŞILAŞTIRMALI
SOLİD ORGAN TRANSPLANTASYONLARINDA İMMÜN MONİTORİZASYON
 SOLİD ORGAN TRANSPLANTASYONLARINDA İMMÜN MONİTORİZASYON Ali ŞENGÜL MEDICALPARK ANTALYA HASTANE KOMPLEKSİ İMMÜNOLOJİ BÖLÜMÜ Organ nakli umudu Beklenen Başarılı Operasyonlar Hayaller ve Komplikasyonlar?
SOLİD ORGAN TRANSPLANTASYONLARINDA İMMÜN MONİTORİZASYON Ali ŞENGÜL MEDICALPARK ANTALYA HASTANE KOMPLEKSİ İMMÜNOLOJİ BÖLÜMÜ Organ nakli umudu Beklenen Başarılı Operasyonlar Hayaller ve Komplikasyonlar?
T.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DÜZLEMSEL HOMOTETİK HAREKETLER ALTINDAT.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
 T.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DÜZLEMSEL HOMOTETİK HAREKETLER ALTINDAT.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ İMMUNOSTİMULAN POLİMERLER İLE ATTENÜE EDİLMİŞ LEISMANIA
T.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DÜZLEMSEL HOMOTETİK HAREKETLER ALTINDAT.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ İMMUNOSTİMULAN POLİMERLER İLE ATTENÜE EDİLMİŞ LEISMANIA
Kanın Bileşenleri. Total kan Miktarı: Vücut Ağırlığı x0.08. Plazma :%55 Hücreler : %45. Plazmanın %90 su
 KAN DOKUSU Kanın Bileşenleri Total kan Miktarı: Vücut Ağırlığı x0.08 Plazma :%55 Hücreler : %45 Plazmanın %90 su Kan Hücreleri Eritrosit Lökosit Trombosit Agranulosit Lenfosit Monosit Granulosit Nötrofil
KAN DOKUSU Kanın Bileşenleri Total kan Miktarı: Vücut Ağırlığı x0.08 Plazma :%55 Hücreler : %45 Plazmanın %90 su Kan Hücreleri Eritrosit Lökosit Trombosit Agranulosit Lenfosit Monosit Granulosit Nötrofil
HAYVANSAL HÜCRELER VE İŞLEVLERİ. YRD. DOÇ. DR. ASLI SADE MEMİŞOĞLU RESİM İŞ ZEMİN KAT ODA: 111
 HAYVANSAL HÜCRELER VE İŞLEVLERİ YRD. DOÇ. DR. ASLI SADE MEMİŞOĞLU RESİM İŞ ZEMİN KAT ODA: 111 asli.memisoglu@deu.edu.tr KONULAR HAYVAN HÜCRESİ HAYVAN, BİTKİ, MANTAR, BAKTERİ HÜCRE FARKLARI HÜCRE ORGANELLERİ
HAYVANSAL HÜCRELER VE İŞLEVLERİ YRD. DOÇ. DR. ASLI SADE MEMİŞOĞLU RESİM İŞ ZEMİN KAT ODA: 111 asli.memisoglu@deu.edu.tr KONULAR HAYVAN HÜCRESİ HAYVAN, BİTKİ, MANTAR, BAKTERİ HÜCRE FARKLARI HÜCRE ORGANELLERİ
Hücre. 1 µm = 0,001 mm (1000 µm = 1 mm)!
!") HÜCRE FİZYOLOJİSİ Hücre Hücre: Tüm canlıların en küçük yapısal ve fonksiyonel ünitesi İnsan vücudunda trilyonlarca hücre bulunur Fare, insan veya filin hücreleri yaklaşık aynı büyüklükte Vücudun büyüklüğü
HÜCRE FİZYOLOJİSİ Hücre Hücre: Tüm canlıların en küçük yapısal ve fonksiyonel ünitesi İnsan vücudunda trilyonlarca hücre bulunur Fare, insan veya filin hücreleri yaklaşık aynı büyüklükte Vücudun büyüklüğü
15- RADYASYONUN NÜKLEİK ASİTLER VE PROTEİNLERE ETKİLERİ
 15- RADYASYONUN NÜKLEİK ASİTLER VE PROTEİNLERE ETKİLERİ İyonlaştırıcı radyasyonların biyomoleküllere örneğin nükleik asitler ve proteinlere olan etkisi hakkında yeterli bilgi yoktur. Ancak, nükleik asitlerden
15- RADYASYONUN NÜKLEİK ASİTLER VE PROTEİNLERE ETKİLERİ İyonlaştırıcı radyasyonların biyomoleküllere örneğin nükleik asitler ve proteinlere olan etkisi hakkında yeterli bilgi yoktur. Ancak, nükleik asitlerden
Su Çiçeği. Suçiçeği Nedir?
 Suçiçeği Nedir? Su çiçeği varisella zoster adı verilen bir virüs tarafından meydana getirilen ateşli bir enfeksiyon hastalığıdır. Varisella zoster virüsü havada 1-2 saat canlı kalan ve çok hızlı çoğalan
Suçiçeği Nedir? Su çiçeği varisella zoster adı verilen bir virüs tarafından meydana getirilen ateşli bir enfeksiyon hastalığıdır. Varisella zoster virüsü havada 1-2 saat canlı kalan ve çok hızlı çoğalan
Bacillus anthracis. Hayvanlarda şarbon etkenidir. Bacillus anthracis. Gram boyama. Bacillus anthracis. Bacillus anthracis
 Bacillus anthracis Gram pozitif, obligat aerop sporlu, çomak şeklinde bakterilerdir. 1µm eninde, 2-4 µm uzunluğunda, konkav sonlanan, kirpiksiz bakterilerdir. Bacillus anthracis in doğal yaşam ortamı topraktır.
Bacillus anthracis Gram pozitif, obligat aerop sporlu, çomak şeklinde bakterilerdir. 1µm eninde, 2-4 µm uzunluğunda, konkav sonlanan, kirpiksiz bakterilerdir. Bacillus anthracis in doğal yaşam ortamı topraktır.
YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
 YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ GÜMÜŞ (Ag) VE TİTANYUM DİOKSİT (TiO 2 ) NANOPARTİKÜLLERİNİN KÜTANÖZ LEISHMANIASIS ETKENİ L.TROPİCA PARAZİTLERİ ÜZERİNDEKİ ANTİLEISHMANIAL ETKİLERİNİN
YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ GÜMÜŞ (Ag) VE TİTANYUM DİOKSİT (TiO 2 ) NANOPARTİKÜLLERİNİN KÜTANÖZ LEISHMANIASIS ETKENİ L.TROPİCA PARAZİTLERİ ÜZERİNDEKİ ANTİLEISHMANIAL ETKİLERİNİN
ORGANİZMALARDA BAĞIŞIKLIK MEKANİZMALARI
 ORGANİZMALARDA BAĞIŞIKLIK MEKANİZMALARI Organizmalarda daha öncede belirtildiği gibi hücresel ve humoral bağışıklık bağışıklık reaksiyonları vardır. Bunlara ilave olarak immünoljik tolerans adı verilen
ORGANİZMALARDA BAĞIŞIKLIK MEKANİZMALARI Organizmalarda daha öncede belirtildiği gibi hücresel ve humoral bağışıklık bağışıklık reaksiyonları vardır. Bunlara ilave olarak immünoljik tolerans adı verilen
Hücre Biyoloji Laboratuarı Güz dönemi Alıştırma Soruları (Dr.Selcen Çelik)
") Hücre Biyoloji Laboratuarı 2014-2015 Güz dönemi Alıştırma Soruları (Dr.Selcen Çelik Konular: ph ve tamponlar, hücre kültür tekniği, mikrometrik ölçüm ph ve Tamponlar 1. ph sı 8.2 olan 500 ml. 20mM Tris/HCl
Hücre Biyoloji Laboratuarı 2014-2015 Güz dönemi Alıştırma Soruları (Dr.Selcen Çelik Konular: ph ve tamponlar, hücre kültür tekniği, mikrometrik ölçüm ph ve Tamponlar 1. ph sı 8.2 olan 500 ml. 20mM Tris/HCl
Hücre Proliferasyonu ve Testleri
 1 Hücre Proliferasyonu ve Testleri Normal Hücre Çoğalması Normal dokularda, hücre bölünmesi ve çoğalması organizmanın devamlılığı için bir gereklilik;r. Hücre çoğalmasının olması gerekenden farklı olması
1 Hücre Proliferasyonu ve Testleri Normal Hücre Çoğalması Normal dokularda, hücre bölünmesi ve çoğalması organizmanın devamlılığı için bir gereklilik;r. Hücre çoğalmasının olması gerekenden farklı olması
BÖLÜM I HÜCRE FİZYOLOJİSİ...
 BÖLÜM I HÜCRE FİZYOLOJİSİ... 1 Bilinmesi Gereken Kavramlar... 1 Giriş... 2 Hücrelerin Fonksiyonel Özellikleri... 2 Hücrenin Kimyasal Yapısı... 2 Hücrenin Fiziksel Yapısı... 4 Hücrenin Bileşenleri... 4
BÖLÜM I HÜCRE FİZYOLOJİSİ... 1 Bilinmesi Gereken Kavramlar... 1 Giriş... 2 Hücrelerin Fonksiyonel Özellikleri... 2 Hücrenin Kimyasal Yapısı... 2 Hücrenin Fiziksel Yapısı... 4 Hücrenin Bileşenleri... 4
İMMUNİZASYON. Bir bireye bağışıklık kazandırma! Bireyin yaşı? İmmunolojik olarak erişkin mi? Maternal antikor? Konak antijene duyarlı mı? Sağlıklı mı?
 İMMUNİZASYON Bir bireye bağışıklık kazandırma! Bireyin yaşı? İmmunolojik olarak erişkin mi? Maternal antikor? Konak antijene duyarlı mı? Sağlıklı mı? Canlıya antijen verdikten belli bir süre sonra, o canlıda
İMMUNİZASYON Bir bireye bağışıklık kazandırma! Bireyin yaşı? İmmunolojik olarak erişkin mi? Maternal antikor? Konak antijene duyarlı mı? Sağlıklı mı? Canlıya antijen verdikten belli bir süre sonra, o canlıda
I- Doğal-doğuştan (innate)var olan bağışıklık
var olan bağışıklık") I- Doğal-doğuştan (innate)var olan bağışıklık Fagositik hücreler (makrofajlar, mast hücreleri) Kompleman sistemi(direkt bakteri hücre membranı parçalayarak diğer immün sistem hücrelerin bunlara atak yapmasına
I- Doğal-doğuştan (innate)var olan bağışıklık Fagositik hücreler (makrofajlar, mast hücreleri) Kompleman sistemi(direkt bakteri hücre membranı parçalayarak diğer immün sistem hücrelerin bunlara atak yapmasına
ADIM ADIM YGS-LYS 55. ADIM CANLILARIN SINIFLANDIRILMASI-15 VİRÜSLER
 ADIM ADIM YGS-LYS 55. ADIM CANLILARIN SINIFLANDIRILMASI-15 VİRÜSLER Virüsler Hücresel yapı da dahil olmak üzere canlıların ortak özelliklerini göstermeyen canlılardır. Prokaryotlardan daha küçüklerdir.
ADIM ADIM YGS-LYS 55. ADIM CANLILARIN SINIFLANDIRILMASI-15 VİRÜSLER Virüsler Hücresel yapı da dahil olmak üzere canlıların ortak özelliklerini göstermeyen canlılardır. Prokaryotlardan daha küçüklerdir.
VİRUS HASTALIKLARINDA TANI YÖNTEMLERİ
 VİRUS HASTALIKLARINDA TANI YÖNTEMLERİ Doç. Dr. Koray Ergünay MD PhD Hacettepe Üniversitesi Tıp Fakültesi Mikrobiyoloji ve Klinik Mikrobiyoloji Anabilim Dalı, Viroloji Ünitesi Viral Enfeksiyonlar... Klinik
VİRUS HASTALIKLARINDA TANI YÖNTEMLERİ Doç. Dr. Koray Ergünay MD PhD Hacettepe Üniversitesi Tıp Fakültesi Mikrobiyoloji ve Klinik Mikrobiyoloji Anabilim Dalı, Viroloji Ünitesi Viral Enfeksiyonlar... Klinik
b. Amaç: Bakterilerin patojenitesine karşı konakçının nasıl cevap verdiği ve savunma mekanizmaları ile ilgili genel bilgi öğretilmesi amaçlanmıştır.
 İMMÜNOLOJİİ I-DERS TANIMLARI 1- Tanım: Konakçı savunma mekanizmalarının öğretilmesi. b. Amaç: Bakterilerin patojenitesine karşı konakçının nasıl cevap verdiği ve savunma mekanizmaları ile ilgili genel
İMMÜNOLOJİİ I-DERS TANIMLARI 1- Tanım: Konakçı savunma mekanizmalarının öğretilmesi. b. Amaç: Bakterilerin patojenitesine karşı konakçının nasıl cevap verdiği ve savunma mekanizmaları ile ilgili genel
İ. Ü İstanbul Tıp Fakültesi Tıbbi Biyoloji Anabilim Dalı Prof. Dr. Filiz Aydın
 İ. Ü İstanbul Tıp Fakültesi Tıbbi Biyoloji Anabilim Dalı Prof. Dr. Filiz Aydın Canlının en küçük birimi Hücre 1665 yılında ilk olarak İngiliz bilim adamı Robert Hooke şişe mantarından kesit alıp mikroskopta
İ. Ü İstanbul Tıp Fakültesi Tıbbi Biyoloji Anabilim Dalı Prof. Dr. Filiz Aydın Canlının en küçük birimi Hücre 1665 yılında ilk olarak İngiliz bilim adamı Robert Hooke şişe mantarından kesit alıp mikroskopta
LENFOİD SİSTEM DR GÖKSAL KESKİN ARALIK-2014
 LENFOİD SİSTEM DR GÖKSAL KESKİN ARALIK-2014 Lenfoid Sistem Lenfositlerin, mononükleer fagositlerin ve diğer yardımcı rol oynayan hücrelerin bulunduğu, yabancı antijenlerin taşınıp yoğunlaştırıldığı, Antijenin
LENFOİD SİSTEM DR GÖKSAL KESKİN ARALIK-2014 Lenfoid Sistem Lenfositlerin, mononükleer fagositlerin ve diğer yardımcı rol oynayan hücrelerin bulunduğu, yabancı antijenlerin taşınıp yoğunlaştırıldığı, Antijenin
Sivrisinek ve Phlebotomus mücadelesinde veya parazit hastalıkların anlatılmasında kullanılan ve de pek anlaşılmayan iki kavram vardır.
 Sivrisinek ve Phlebotomus mücadelesinde veya parazit hastalıkların anlatılmasında kullanılan ve de pek anlaşılmayan iki kavram vardır. Bu kavramlar ; vektör ve konak dır. VEKTÖR NEDİR? Herhangi bir bulaşıcı
Sivrisinek ve Phlebotomus mücadelesinde veya parazit hastalıkların anlatılmasında kullanılan ve de pek anlaşılmayan iki kavram vardır. Bu kavramlar ; vektör ve konak dır. VEKTÖR NEDİR? Herhangi bir bulaşıcı
İSTANBUL MEDENİYET ÜNİVERSİTESİ TIP FAKÜLTESİ TIBBİ MİKROBİYOLOJİ ANABİLİM DALI MEZUNİYET SONRASI (UZMANLIK) EĞİTİMİ DERS MÜFREDATI
 EĞİTİMİ DERS MÜFREDATI") İSTANBUL MEDENİYET ÜNİVERSİTESİ TIP FAKÜLTESİ TIBBİ MİKROBİYOLOJİ ANABİLİM DALI MEZUNİYET SONRASI (UZMANLIK) EĞİTİMİ DERS MÜFREDATI DERS KODU ve ADI TMİK 001: Vaka Değerlendirme Toplantısı TMİK 002: Makale
İSTANBUL MEDENİYET ÜNİVERSİTESİ TIP FAKÜLTESİ TIBBİ MİKROBİYOLOJİ ANABİLİM DALI MEZUNİYET SONRASI (UZMANLIK) EĞİTİMİ DERS MÜFREDATI DERS KODU ve ADI TMİK 001: Vaka Değerlendirme Toplantısı TMİK 002: Makale
LİZOZOMLAR Doç. Dr. Mehmet Güven
 LİZOZOMLAR Doç.. Dr. Mehmet GüvenG Lizozomlar tek bir membran ile çevrili evrili veziküler yapılı organellerdir. Lizozomlar eritrosit dışıd ışındaki tüm t m hayvan hücrelerinde h bulunur. Ortalama olarak
LİZOZOMLAR Doç.. Dr. Mehmet GüvenG Lizozomlar tek bir membran ile çevrili evrili veziküler yapılı organellerdir. Lizozomlar eritrosit dışıd ışındaki tüm t m hayvan hücrelerinde h bulunur. Ortalama olarak
Staphylococcus Gram pozitif koklardır.
 Staphylococcus Gram pozitif koklardır. 0.8-1µm çapında küçük, yuvarlak veya oval bakterilerdir. Hareketsizdirler. Spor oluşturmazlar ve katalaz enzimi üretirler. Gram boyama Koagülaz, alfatoksin, lökosidin,
Staphylococcus Gram pozitif koklardır. 0.8-1µm çapında küçük, yuvarlak veya oval bakterilerdir. Hareketsizdirler. Spor oluşturmazlar ve katalaz enzimi üretirler. Gram boyama Koagülaz, alfatoksin, lökosidin,
ORTOPEDİK PROTEZ ENFEKSİYONLARINDA SONİKASYON DENEYİMİ
 ORTOPEDİK PROTEZ ENFEKSİYONLARINDA SONİKASYON DENEYİMİ Dr. Şua Sümer Selçuk Üniversitesi Tıp Fakültesi Enf. Hast. ve Klin. Mikr. AD 17 Mayıs 2016 Prostetik eklem ameliyatları yaşlı popülasyonun artışına
ORTOPEDİK PROTEZ ENFEKSİYONLARINDA SONİKASYON DENEYİMİ Dr. Şua Sümer Selçuk Üniversitesi Tıp Fakültesi Enf. Hast. ve Klin. Mikr. AD 17 Mayıs 2016 Prostetik eklem ameliyatları yaşlı popülasyonun artışına
Yasemin Budama Kılınç1, Rabia Çakır Koç1, Sevim Meşe2, Selim Badur2,3
 Yasemin Budama Kılınç1, Rabia Çakır Koç1, Sevim Meşe2, Selim Badur2,3 1 Yıldız Teknik Üniversitesi, Biyomühendislik Bölümü, 34220, İstanbul 2 İstanbul Üniversitesi, İst. Tıp Fak., Mikrobiyoloji ABD, Viroloji
Yasemin Budama Kılınç1, Rabia Çakır Koç1, Sevim Meşe2, Selim Badur2,3 1 Yıldız Teknik Üniversitesi, Biyomühendislik Bölümü, 34220, İstanbul 2 İstanbul Üniversitesi, İst. Tıp Fak., Mikrobiyoloji ABD, Viroloji
ÇEKİRDEK EĞİTİM PROGRAMI
 ÇEKİRDEK EĞİTİM PROGRAMI Tıp Fakülteleri Mezuniyet Öncesi İmmünoloji Eğitim Programı Önerisi in hücre ve dokuları ilgi hücrelerini isim ve işlevleri ile bilir. Kemik iliği, lenf nodu, ve dalağın anatomisi,
ÇEKİRDEK EĞİTİM PROGRAMI Tıp Fakülteleri Mezuniyet Öncesi İmmünoloji Eğitim Programı Önerisi in hücre ve dokuları ilgi hücrelerini isim ve işlevleri ile bilir. Kemik iliği, lenf nodu, ve dalağın anatomisi,
Moleküler Yöntemlerin Klinik Mikrobiyolojide Kullanımı Ne zaman? Nerede? Ne kadar? Klinik Parazitoloji
 Moleküler Yöntemlerin Klinik Mikrobiyolojide Kullanımı Ne zaman? Nerede? Ne kadar? Klinik Parazitoloji Metin Korkmaz Ege Üniversitesi Tıp Fakültesi Tıbbi Parazitoloji AD İnsandaki Paraziter Hastalıkların
Moleküler Yöntemlerin Klinik Mikrobiyolojide Kullanımı Ne zaman? Nerede? Ne kadar? Klinik Parazitoloji Metin Korkmaz Ege Üniversitesi Tıp Fakültesi Tıbbi Parazitoloji AD İnsandaki Paraziter Hastalıkların
MİKROBİYOLOJİ SORU KAMPI 2015
 Canlıların prokaryot ve ökoaryot olma özelliğini hücre komponentlerinden hangisi belirler? MİKROBİYOLOJİ SORU KAMPI 2015 B. Stoplazmik membran C. Golgi membranı D. Nükleer membran E. Endoplazmik retikulum
Canlıların prokaryot ve ökoaryot olma özelliğini hücre komponentlerinden hangisi belirler? MİKROBİYOLOJİ SORU KAMPI 2015 B. Stoplazmik membran C. Golgi membranı D. Nükleer membran E. Endoplazmik retikulum
HÜCRE ZARINDA TAŞIMA PROF. DR. SERKAN YILMAZ
 HÜCRE ZARINDA TAŞIMA PROF. DR. SERKAN YILMAZ Hücre içi ve hücre dışı sıvılar bileşimleri yönünden oldukça farklıdır. Hücre içi sıvı intraselüler sıvı, hücre dışı sıvı ise ekstraselüler sıvı adını alır.
HÜCRE ZARINDA TAŞIMA PROF. DR. SERKAN YILMAZ Hücre içi ve hücre dışı sıvılar bileşimleri yönünden oldukça farklıdır. Hücre içi sıvı intraselüler sıvı, hücre dışı sıvı ise ekstraselüler sıvı adını alır.
TEMEL ARAŞTIRMA TEKNİKLERİ II
 TEMEL ARAŞTIRMA TEKNİKLERİ II 6,7,8 HAFTA Prof. Dr. Eser ELÇİN Hücre kültüründe temel işlemler: besiyeri hazırlama, hücre ekimi, besiyeri değiştirme, alt kültüre geçiş, hücre sayımı, pasajlama, kontaminasyon
TEMEL ARAŞTIRMA TEKNİKLERİ II 6,7,8 HAFTA Prof. Dr. Eser ELÇİN Hücre kültüründe temel işlemler: besiyeri hazırlama, hücre ekimi, besiyeri değiştirme, alt kültüre geçiş, hücre sayımı, pasajlama, kontaminasyon
Hümoral İmmün Yanıt ve Antikorlar
 Hümoral İmmün Yanıt ve Antikorlar H. Barbaros ORAL Uludağ Üniversitesi Tıp Fakültesi İmmünoloji Anabilim Dalı Edinsel immün sistemin antijenleri bağlamak için kullandığı 3 molekül sınıfı: I.Antikorlar,
Hümoral İmmün Yanıt ve Antikorlar H. Barbaros ORAL Uludağ Üniversitesi Tıp Fakültesi İmmünoloji Anabilim Dalı Edinsel immün sistemin antijenleri bağlamak için kullandığı 3 molekül sınıfı: I.Antikorlar,
Mycobacterium. Mycobacterium hücre duvarının lipid içeriği oldukça fazladır ve mikolik asit içerir
 Mycobacterium Mycobacteriaceae ailesi üyeleri uzun, ince, çomak şekilli, hareketsiz bakterilerdir. Özel ayırt edici boyalarla bir kez boyandıklarında seyreltik asitlerle boyayı vermemeleri yani dekolorize
Mycobacterium Mycobacteriaceae ailesi üyeleri uzun, ince, çomak şekilli, hareketsiz bakterilerdir. Özel ayırt edici boyalarla bir kez boyandıklarında seyreltik asitlerle boyayı vermemeleri yani dekolorize
Tam Kan Analizi. Yrd.Doç.Dr.Filiz BAKAR ATEŞ
 Tam Kan Analizi Yrd.Doç.Dr.Filiz BAKAR ATEŞ Tam Kan Analizi Tam kan analizi, en sık kullanılan kan testlerinden biridir. Kandaki 3 major hücreyi analiz eder: 1. Eritrositler 2. Lökositler 3. Plateletler
Tam Kan Analizi Yrd.Doç.Dr.Filiz BAKAR ATEŞ Tam Kan Analizi Tam kan analizi, en sık kullanılan kan testlerinden biridir. Kandaki 3 major hücreyi analiz eder: 1. Eritrositler 2. Lökositler 3. Plateletler
VİROLOJİ -I Antiviral İmmunite
 VİROLOJİ -I Antiviral İmmunite Prof.Dr. Yılmaz Akça Prof.Dr. Feray Alkan Prof.Dr. Aykut Özkul Prof. Dr. Seval Bilge-Dağalp Prof.Dr. M. Taner Karaoğlu Prof.Dr. Tuba Çiğdem Oğuzoğlu DOĞAL SAVUNMA HATLARI-DOĞAL
VİROLOJİ -I Antiviral İmmunite Prof.Dr. Yılmaz Akça Prof.Dr. Feray Alkan Prof.Dr. Aykut Özkul Prof. Dr. Seval Bilge-Dağalp Prof.Dr. M. Taner Karaoğlu Prof.Dr. Tuba Çiğdem Oğuzoğlu DOĞAL SAVUNMA HATLARI-DOĞAL
EĞİTİM-ÖĞRETİM BAHAR YARI YILI LİSANSÜSTÜ DERS PROGRAMI. Tarih Saat Konu Yer Anlatan
 2016- EĞİTİM-ÖĞRETİM BAHAR YARI YILI LİSANSÜSTÜ DERS PROGRAMI EĞİTİM-ÖĞRETİM BAŞLANGIÇ: 13 Şubat EĞİTİM-ÖĞRETİM BİTİŞ: 18 Mayıs Bitirme sınav tarihleri : 22 Mayıs-2 Haziran Bütünleme sınav tarihleri: 12-23
2016- EĞİTİM-ÖĞRETİM BAHAR YARI YILI LİSANSÜSTÜ DERS PROGRAMI EĞİTİM-ÖĞRETİM BAŞLANGIÇ: 13 Şubat EĞİTİM-ÖĞRETİM BİTİŞ: 18 Mayıs Bitirme sınav tarihleri : 22 Mayıs-2 Haziran Bütünleme sınav tarihleri: 12-23
Doç. Dr. Fatih ÇALIŞKAN Sakarya Üniversitesi, Teknoloji Fak. Metalurji ve Malzeme Mühendisliği EABD
 BİYOUYUMLULUK (BIO-COMPATIBILITY) 10993-1 Bir materyalin biyo-uyumluluğunun test edilmesi için gerekli testlerin tümünü içerir. (Toksisite, Hemoliz, sitotoksisite, sistemik toksisite,...vs.) Hammaddelerin
BİYOUYUMLULUK (BIO-COMPATIBILITY) 10993-1 Bir materyalin biyo-uyumluluğunun test edilmesi için gerekli testlerin tümünü içerir. (Toksisite, Hemoliz, sitotoksisite, sistemik toksisite,...vs.) Hammaddelerin
DİCLE ÜNİVERSİTESİ TIP FAKÜLTESİ DÖNEM II. KAN-DOLAŞIM ve SOLUNUM DERS KURULU
 DİCLE ÜNİVERSİTESİ TIP FAKÜLTESİ DÖNEM II KAN-DOLAŞIM ve SOLUNUM DERS KURULU Doç.Dr. Engin DEVECİ İMMÜN SİSTEM TİPLERİ I- Doğal-doğuştan (innate)var olan bağışıklık Fagositik hücreler (makrofajlar, mast
DİCLE ÜNİVERSİTESİ TIP FAKÜLTESİ DÖNEM II KAN-DOLAŞIM ve SOLUNUM DERS KURULU Doç.Dr. Engin DEVECİ İMMÜN SİSTEM TİPLERİ I- Doğal-doğuştan (innate)var olan bağışıklık Fagositik hücreler (makrofajlar, mast
HÜCRESEL İMMÜNİTENİN EFEKTÖR MEKANİZMALARI. Hücre İçi Mikropların Yok Edilmesi
 HÜCRESEL İMMÜNİTENİN EFEKTÖR MEKANİZMALARI Hücre İçi Mikropların Yok Edilmesi Hücre içi mikropları yok etmekle görevli özelleşmiş immün mekanizmalar hücre aracılı immüniteyi oluştururlar. Hücresel immünitenin
HÜCRESEL İMMÜNİTENİN EFEKTÖR MEKANİZMALARI Hücre İçi Mikropların Yok Edilmesi Hücre içi mikropları yok etmekle görevli özelleşmiş immün mekanizmalar hücre aracılı immüniteyi oluştururlar. Hücresel immünitenin
İÇİNDEKİLER BÖLÜM 1: MİKROBİYOLOJİYE GİRİŞ...1 BÖLÜM 2: MİKROORGANİZMALARIN MORFOLOJİLERİ.13 BÖLÜM 3: MİKROORGANİZMALARIN HÜCRE YAPILARI...
 İÇİNDEKİLER BÖLÜM 1: MİKROBİYOLOJİYE GİRİŞ...1 1.1. Tanım ve Kapsam...1 1.2. Mikrobiyoloji Biliminin Gelişmesi...2 1.3. Mikroorganizmaların Hayatımızdaki Önemi...5 1.3.1. Mikroorganizmaların Yararları...5
İÇİNDEKİLER BÖLÜM 1: MİKROBİYOLOJİYE GİRİŞ...1 1.1. Tanım ve Kapsam...1 1.2. Mikrobiyoloji Biliminin Gelişmesi...2 1.3. Mikroorganizmaların Hayatımızdaki Önemi...5 1.3.1. Mikroorganizmaların Yararları...5
DEHİDRE KÖPEKLERDE BİKARBONATLI SODYUM KLORÜR SOLÜSYONUNUN HEMATOLOJİK VE BİYOKİMYASAL PARAMETRELERE ETKİSİ
 T.C. ADNAN MENDERES ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ İÇ HASTALIKLARI ANABİLİM DALI VİH-YL 2007 0002 DEHİDRE KÖPEKLERDE BİKARBONATLI SODYUM KLORÜR SOLÜSYONUNUN HEMATOLOJİK VE BİYOKİMYASAL PARAMETRELERE
T.C. ADNAN MENDERES ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ İÇ HASTALIKLARI ANABİLİM DALI VİH-YL 2007 0002 DEHİDRE KÖPEKLERDE BİKARBONATLI SODYUM KLORÜR SOLÜSYONUNUN HEMATOLOJİK VE BİYOKİMYASAL PARAMETRELERE
2. HAFTA MİKROSKOPLAR
 2. HAFTA MİKROSKOPLAR MİKROSKOPLAR Hücreler çok küçük olduğundan (3-200 µm) mikroskop kullanılması zorunludur. Soğan zarı, parmak arası zarlar gibi çok ince yapılar, kesit almadan ve mikroskopsuz incelenebilir.
2. HAFTA MİKROSKOPLAR MİKROSKOPLAR Hücreler çok küçük olduğundan (3-200 µm) mikroskop kullanılması zorunludur. Soğan zarı, parmak arası zarlar gibi çok ince yapılar, kesit almadan ve mikroskopsuz incelenebilir.
Deneysel Hayvan Modelinde Candida Tropicalis Peritonitinin Tedavisinde Kaspofungin ve Amfoterisin B Etkinliğinin Karşılaştırılması
 Deneysel Hayvan Modelinde Candida Tropicalis Peritonitinin Tedavisinde Kaspofungin ve Amfoterisin B Etkinliğinin Karşılaştırılması Melis Demirci, Özlem Tünger, Kenan Değerli, Şebnem Şenol, Çiğdem Banu
Deneysel Hayvan Modelinde Candida Tropicalis Peritonitinin Tedavisinde Kaspofungin ve Amfoterisin B Etkinliğinin Karşılaştırılması Melis Demirci, Özlem Tünger, Kenan Değerli, Şebnem Şenol, Çiğdem Banu
BİYOKİMYA ANABİLİM DALI LİSANSÜSTÜ DERS PROGRAMI
 BİYOKİMYA ANABİLİM DALI LİSANSÜSTÜ DERS PROGRAMI SAĞLIK BİLİMLERİ ENSİTÜSÜ İ Yüksek Lisans Programı SZR 101 Bilimsel Araştırma Ders (T+ U) 2+2 3 6 AD SZR 103 Akılcı İlaç Kullanımı 2+0 2 5 Enstitünün Belirlediği
BİYOKİMYA ANABİLİM DALI LİSANSÜSTÜ DERS PROGRAMI SAĞLIK BİLİMLERİ ENSİTÜSÜ İ Yüksek Lisans Programı SZR 101 Bilimsel Araştırma Ders (T+ U) 2+2 3 6 AD SZR 103 Akılcı İlaç Kullanımı 2+0 2 5 Enstitünün Belirlediği
KAN DOKUSU. Prof. Dr. Levent ERGÜN
 KAN DOKUSU Prof. Dr. Levent ERGÜN 1 Kan Dokusu Plazma (sıvı) ve şekilli elemanlarından oluşur Plazma fundememtal substans olarak kabul edilir. Kanın fonksiyonları Transport Gaz, besin, hormon, atık maddeler,
KAN DOKUSU Prof. Dr. Levent ERGÜN 1 Kan Dokusu Plazma (sıvı) ve şekilli elemanlarından oluşur Plazma fundememtal substans olarak kabul edilir. Kanın fonksiyonları Transport Gaz, besin, hormon, atık maddeler,
Edinsel İmmün Yanıt Güher Saruhan- Direskeneli
 Edinsel İmmün Yanıt Güher Saruhan- Direskeneli İTF Fizyoloji AD Doğal bağışıklık Edinsel bağışıklık Hızlı yanıt (saatler) Sabit R yapıları Sınırlı çeşidi tanıma Yanıt sırasında değişmez Yavaş yanıt (Gün-hafta)
Edinsel İmmün Yanıt Güher Saruhan- Direskeneli İTF Fizyoloji AD Doğal bağışıklık Edinsel bağışıklık Hızlı yanıt (saatler) Sabit R yapıları Sınırlı çeşidi tanıma Yanıt sırasında değişmez Yavaş yanıt (Gün-hafta)
Protein Ekstraksiyonu
 Protein Ekstraksiyonu Dr.Gaye Güler Tezel Hacettepe Üniversitesi Tıp Fakültesi Patoloji Anabilim Dalı Proteinler tüm canlı organizmalar için en önemli makromoleküllerden biridir. Bazıları yapısal komponentleri
Protein Ekstraksiyonu Dr.Gaye Güler Tezel Hacettepe Üniversitesi Tıp Fakültesi Patoloji Anabilim Dalı Proteinler tüm canlı organizmalar için en önemli makromoleküllerden biridir. Bazıları yapısal komponentleri
Kocaeli Üniversitesi Tıp Fakültesi Çocuk Sağlığı ve Hastalıkları Anabilim Dalı Enfeksiyon Hastalıkları BD Olgu Sunumu 13 Ekim 2018 Perşembe
 Kocaeli Üniversitesi Tıp Fakültesi Çocuk Sağlığı ve Hastalıkları Anabilim Dalı Enfeksiyon Hastalıkları BD Olgu Sunumu 13 Ekim 2018 Perşembe Dr. Ayşe Tekin Yılmaz Olgu Sekiz yaş, erkek hasta Yakınma Sırt
Kocaeli Üniversitesi Tıp Fakültesi Çocuk Sağlığı ve Hastalıkları Anabilim Dalı Enfeksiyon Hastalıkları BD Olgu Sunumu 13 Ekim 2018 Perşembe Dr. Ayşe Tekin Yılmaz Olgu Sekiz yaş, erkek hasta Yakınma Sırt
24 Şubat Enflamasyon DETAE Prof. Dr. Günnur Deniz Dr. Sema Bilgiç
 IMMN 7005. İMMÜN SİSTEM MORFOLOJİSİ 24 Şubat Enflamasyon DETAE Prof. Dr. Günnur Deniz Dr. Sema Bilgiç 3 Mart Hormon Etki Mekanizması Biyokimya Prof.Dr.Beyhan Ömer 10 Mart Isı Şoku Proteinleri DETAE Doç.Dr.
IMMN 7005. İMMÜN SİSTEM MORFOLOJİSİ 24 Şubat Enflamasyon DETAE Prof. Dr. Günnur Deniz Dr. Sema Bilgiç 3 Mart Hormon Etki Mekanizması Biyokimya Prof.Dr.Beyhan Ömer 10 Mart Isı Şoku Proteinleri DETAE Doç.Dr.
LÖKOSİT. WBC; White Blood Cell,; Akyuvar. Lökosit için normal değer : Lökosit sayısını arttıran sebepler: Lökosit sayısını azaltan sebepler:
 LÖKOSİT WBC; White Blood Cell,; Akyuvar Lökositler kanın beyaz hücreleridir ve vücudun savunmasında görev alırlar. Lökositler kemik iliğinde yapılır ve kan yoluyla bütün dokulara ulaşır vücudumuzu mikrop
LÖKOSİT WBC; White Blood Cell,; Akyuvar Lökositler kanın beyaz hücreleridir ve vücudun savunmasında görev alırlar. Lökositler kemik iliğinde yapılır ve kan yoluyla bütün dokulara ulaşır vücudumuzu mikrop
1-GİRİ 1.1- BİYOKİMYANIN TANIMI VE KONUSU.-
 1-GİRİ 1.1- BİYOKİMYANIN TANIMI VE KONUSU.- Biyokimya sözcüğü biyolojik kimya (=yaşam kimyası) teriminin kısaltılmış şeklidir. Daha eskilerde, fizyolojik kimya terimi kullanılmıştır. Gerçekten de Biyokimya
1-GİRİ 1.1- BİYOKİMYANIN TANIMI VE KONUSU.- Biyokimya sözcüğü biyolojik kimya (=yaşam kimyası) teriminin kısaltılmış şeklidir. Daha eskilerde, fizyolojik kimya terimi kullanılmıştır. Gerçekten de Biyokimya
Rahim ağzı kanseri hücreleri doku kültürü mikroskopik görüntüsü.
 Doç.Dr.Engin DEVECİ HÜCRE KÜLTÜRÜ Hücre Kültürü Araştırma Laboratuvarı, çeşitli hücrelerin invitro kültürlerini yaparak araştırmacılara kanser, kök hücre, hücre mekaniği çalışmaları gibi konularda hücre
Doç.Dr.Engin DEVECİ HÜCRE KÜLTÜRÜ Hücre Kültürü Araştırma Laboratuvarı, çeşitli hücrelerin invitro kültürlerini yaparak araştırmacılara kanser, kök hücre, hücre mekaniği çalışmaları gibi konularda hücre
VİRAL ENFEKSİYONLAR VE KORUNMA. Yrd. Doç. Dr. Banu KAŞKATEPE
 VİRAL ENFEKSİYONLAR VE KORUNMA Yrd. Doç. Dr. Banu KAŞKATEPE VİRAL HASTALIKLARDA İMMÜNİTE Virüsler konak hücreye girdikten sonra çoğalır ve viral çoğalma belirli bir düzeye ulaştığında hastalık semptomları
VİRAL ENFEKSİYONLAR VE KORUNMA Yrd. Doç. Dr. Banu KAŞKATEPE VİRAL HASTALIKLARDA İMMÜNİTE Virüsler konak hücreye girdikten sonra çoğalır ve viral çoğalma belirli bir düzeye ulaştığında hastalık semptomları
HÜCRENİN YAŞAM DÖNGÜSÜ
 HÜCRENİN YAŞAM DÖNGÜSÜ *Hücrenin yaşam döngüsü: Hücrenin; bir bölünme sonundan, ikinci bir bölünme sonuna kadar olan zaman sürecinde; geçirdiği yaşamsal olaylara hücrenin yaşam döngüsü denir. Hücreler,
HÜCRENİN YAŞAM DÖNGÜSÜ *Hücrenin yaşam döngüsü: Hücrenin; bir bölünme sonundan, ikinci bir bölünme sonuna kadar olan zaman sürecinde; geçirdiği yaşamsal olaylara hücrenin yaşam döngüsü denir. Hücreler,
Kanın fonksiyonel olarak üstlendiği görevler
 EGZERSİZ VE KAN Kanın fonksiyonel olarak üstlendiği görevler Akciğerden dokulara O2 taşınımı, Dokudan akciğere CO2 taşınımı, Sindirim organlarından hücrelere besin maddeleri taşınımı, Hücreden atık maddelerin
EGZERSİZ VE KAN Kanın fonksiyonel olarak üstlendiği görevler Akciğerden dokulara O2 taşınımı, Dokudan akciğere CO2 taşınımı, Sindirim organlarından hücrelere besin maddeleri taşınımı, Hücreden atık maddelerin
CANDİDA İLE UYARILMIŞ VAJİNAL VE BUKKAL EPİTEL HÜCRELERİNİN SİTOKİN ÜRETİMİ
 CANDİDA İLE UYARILMIŞ VAJİNAL VE BUKKAL EPİTEL HÜCRELERİNİN SİTOKİN ÜRETİMİ Emine Yeşilyurt, Sevgi Özyeğen Aslan, Ayşe Kalkancı, Işıl Fidan, Semra Kuştimur Gazi Üniversitesi Tıp Fakültesi Tıbbi Mikrobiyoloji
CANDİDA İLE UYARILMIŞ VAJİNAL VE BUKKAL EPİTEL HÜCRELERİNİN SİTOKİN ÜRETİMİ Emine Yeşilyurt, Sevgi Özyeğen Aslan, Ayşe Kalkancı, Işıl Fidan, Semra Kuştimur Gazi Üniversitesi Tıp Fakültesi Tıbbi Mikrobiyoloji
KANSER AŞILARI. Prof. Dr. Tezer Kutluk Hacettepe Üniversitesi
 KANSER AŞILARI Prof. Dr. Tezer Kutluk Hacettepe Üniversitesi Bir Halk Sağlığı Sorunu Şu an dünyada 24.600.000 kanserli vardır. Her yıl 10.9 milyon kişi kansere yakalanmaktadır. 2020 yılında bu rakam %50
KANSER AŞILARI Prof. Dr. Tezer Kutluk Hacettepe Üniversitesi Bir Halk Sağlığı Sorunu Şu an dünyada 24.600.000 kanserli vardır. Her yıl 10.9 milyon kişi kansere yakalanmaktadır. 2020 yılında bu rakam %50
ANTRAKS (ŞARBON) septisemik, bulaşıcı, zoonoz
 septisemik, bulaşıcı, zoonoz") ANTRAKS (ŞARBON) septisemik, bulaşıcı, zoonoz ÖLÜMden hemen önce ya da sonra doğal boşluklardan KAN PIHTILAŞMAMA KOYU RENK alma DALAKta büyüme ÖDEM ETİYOLOJİ Bacillus anthracis Gram pozitif kapsüllü *
ANTRAKS (ŞARBON) septisemik, bulaşıcı, zoonoz ÖLÜMden hemen önce ya da sonra doğal boşluklardan KAN PIHTILAŞMAMA KOYU RENK alma DALAKta büyüme ÖDEM ETİYOLOJİ Bacillus anthracis Gram pozitif kapsüllü *
Bağışıklık sistemi nasıl çalışır?
 On5yirmi5.com Bağışıklık sistemi nasıl çalışır? İnsanda bağışıklık sistemi, özellik ve görevleri nelerdir? Kaç çeşit bağışıklık sistemi vardır? Yayın Tarihi : 23 Ekim 2012 Salı (oluşturma : 10/3/2017)
On5yirmi5.com Bağışıklık sistemi nasıl çalışır? İnsanda bağışıklık sistemi, özellik ve görevleri nelerdir? Kaç çeşit bağışıklık sistemi vardır? Yayın Tarihi : 23 Ekim 2012 Salı (oluşturma : 10/3/2017)
T.C. KAHRAMANMARAŞ SÜTÇÜ İMAM ÜNİVERSİTESİ TIP FAKÜLTESİ DÖNEM I EĞİTİM - ÖĞRETİM YILI Tıbbi Bilimlere Giriş Ders Kurulu DERS KURULU-I
 T.C. KAHRAMANMARAŞ SÜTÇÜ İMAM ÜNİVERSİTESİ TIP FAKÜLTESİ DÖNEM I 2016-2017 EĞİTİM - ÖĞRETİM YILI Tıbbi Bilimlere Giriş Ders Kurulu DERS KURULU-I 19 EYLÜL 02 ARALIK 2016 (11 HAFTA) DERSLER TEORİK PRATİK
T.C. KAHRAMANMARAŞ SÜTÇÜ İMAM ÜNİVERSİTESİ TIP FAKÜLTESİ DÖNEM I 2016-2017 EĞİTİM - ÖĞRETİM YILI Tıbbi Bilimlere Giriş Ders Kurulu DERS KURULU-I 19 EYLÜL 02 ARALIK 2016 (11 HAFTA) DERSLER TEORİK PRATİK
Doğal Bağışıklık. İnsan doğar doğmaz hazırdır
 Doğal Bağışıklık 1 Doğal Bağışıklık İnsan doğar doğmaz hazırdır 2 Mikrop vücuda girdiği zaman doğal bağışıklık onunla saatler içinde savaşır. 3 Doğal bağışıklık ikinci görev olarak adaptif immün cevabı
Doğal Bağışıklık 1 Doğal Bağışıklık İnsan doğar doğmaz hazırdır 2 Mikrop vücuda girdiği zaman doğal bağışıklık onunla saatler içinde savaşır. 3 Doğal bağışıklık ikinci görev olarak adaptif immün cevabı
HÜCRE DONDURMA VE ÇÖZME. Uzm. Mol. Bio. Gamze ÇAĞATAY
 HÜCRE DONDURMA VE ÇÖZME Uzm. Mol. Bio. Gamze ÇAĞATAY Neden Dondurma? Hücreleri saklamak Düşük pasajlarda araştırma yapmak Hücrenin orijinal yapısını korumak Genetik stabiliteyi korumak Mikrobiyal ve çapraz
HÜCRE DONDURMA VE ÇÖZME Uzm. Mol. Bio. Gamze ÇAĞATAY Neden Dondurma? Hücreleri saklamak Düşük pasajlarda araştırma yapmak Hücrenin orijinal yapısını korumak Genetik stabiliteyi korumak Mikrobiyal ve çapraz
1. ÜNİTE: YAŞAM BİLİMİ BİYOLOJİ...10
 İçindekiler 1. ÜNİTE: YAŞAM BİLİMİ BİYOLOJİ...10 1. BÖLÜM: BİLİMSEL BİLGİNİN DOĞASI ve BİYOLOJİ... 12 A. BİLİMSEL ÇALIŞMA YÖNTEMİ... 12 1. Bilim İnsanı ve Bilim... 12 B. BİLİMSEL YÖNTEMİN AŞAMALARI...
İçindekiler 1. ÜNİTE: YAŞAM BİLİMİ BİYOLOJİ...10 1. BÖLÜM: BİLİMSEL BİLGİNİN DOĞASI ve BİYOLOJİ... 12 A. BİLİMSEL ÇALIŞMA YÖNTEMİ... 12 1. Bilim İnsanı ve Bilim... 12 B. BİLİMSEL YÖNTEMİN AŞAMALARI...
Hücresel İmmünite Dicle Güç
 Hücresel İmmünite Dicle Güç dguc@hacettepe.edu.tr kekik imus Kalbe yakınlığı ve Esrarengiz hale Ruhun oturduğu yer Ruh cesaret yiğitlik Yunanlı Hekim MS 1.yy Kalp, pankreas, timus imus yaşla küçülür (timik
Hücresel İmmünite Dicle Güç dguc@hacettepe.edu.tr kekik imus Kalbe yakınlığı ve Esrarengiz hale Ruhun oturduğu yer Ruh cesaret yiğitlik Yunanlı Hekim MS 1.yy Kalp, pankreas, timus imus yaşla küçülür (timik
T.C. MUSTAFA KEMAL ÜNİVERSİTESİ TAYFUR ATA SÖKMEN TIP FAKÜLTESİ EĞİTİM - ÖĞRETİM YILI DÖNEM II VI. DERS KURULU (4 HAFTA)
") T.C. MUSTAFA KEMAL ÜNİVERSİTESİ TAYFUR ATA SÖKMEN TIP FAKÜLTESİ 2016-2017 EĞİTİM - ÖĞRETİM YILI DÖNEM II VI. DERS KURULU (4 HAFTA) 1901206 HASTALIKLARIN BİYOLOJİK TEMELİ DERS KURULU-I DEKAN DEKAN YRD.
T.C. MUSTAFA KEMAL ÜNİVERSİTESİ TAYFUR ATA SÖKMEN TIP FAKÜLTESİ 2016-2017 EĞİTİM - ÖĞRETİM YILI DÖNEM II VI. DERS KURULU (4 HAFTA) 1901206 HASTALIKLARIN BİYOLOJİK TEMELİ DERS KURULU-I DEKAN DEKAN YRD.
İNÖNÜ ÜNİVERSİTESİ SAĞLIK HİZMETLERİ MESLEK YÜKSEKOKULU TIBBİ LABORATUVAR TEKNİKLERİ PROGRAMI II DERS İÇERİKLERİ:
 İNÖNÜ ÜNİVERSİTESİ SAĞLIK HİZMETLERİ MESLEK YÜKSEKOKULU TIBBİ LABORATUVAR TEKNİKLERİ PROGRAMI II DERS İÇERİKLERİ: HEMATOLOJİ (2 4 6) 1 Hematolojiye giriş-tanımı, Hastaların genel değerlendirilmesi, 2 Kan,
İNÖNÜ ÜNİVERSİTESİ SAĞLIK HİZMETLERİ MESLEK YÜKSEKOKULU TIBBİ LABORATUVAR TEKNİKLERİ PROGRAMI II DERS İÇERİKLERİ: HEMATOLOJİ (2 4 6) 1 Hematolojiye giriş-tanımı, Hastaların genel değerlendirilmesi, 2 Kan,
Hücre Nükleusu, Nükleus Membranı, Nükleus Porları. Doç. Dr. Ahmet Özaydın
 Hücre Nükleusu, Nükleus Membranı, Nükleus Porları Doç. Dr. Ahmet Özaydın Nükleus (çekirdek) ökaryotlar ile prokaryotları ayıran temel özelliktir. Çekirdek hem genetik bilginin deposu hem de kontrol merkezidir.
Hücre Nükleusu, Nükleus Membranı, Nükleus Porları Doç. Dr. Ahmet Özaydın Nükleus (çekirdek) ökaryotlar ile prokaryotları ayıran temel özelliktir. Çekirdek hem genetik bilginin deposu hem de kontrol merkezidir.
(ZORUNLU) MOLEKÜLER İMMÜNOLOJİ I (TBG 607 TEORİK 3, 3 KREDİ)
 MOLEKÜLER İMMÜNOLOJİ I (TBG 607 TEORİK 3, 3 KREDİ)") T. C. İSTANBUL BİLİM ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ VE GENETİK ANABİLİM DALI YÜKSEK LİSANS PROGRAMI 2015-2016 EĞİTİM-ÖĞRETİM YILI DERS İÇERİKLERİ I. YARIYIL (ZORUNLU) MOLEKÜLER
T. C. İSTANBUL BİLİM ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ VE GENETİK ANABİLİM DALI YÜKSEK LİSANS PROGRAMI 2015-2016 EĞİTİM-ÖĞRETİM YILI DERS İÇERİKLERİ I. YARIYIL (ZORUNLU) MOLEKÜLER
T.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
 T.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ LEİSHMANİA PARAZİTLERİNDEN LİPOFOSFOGLİKAN (LPG) SAFLAŞTIRILMASI VE AŞI MODELİ OLARAK LPG-POLİMER KONJUGATININ ANTİLEİSHMANİAL ETKİNLİĞİNİN IN VITRO
T.C. YILDIZ TEKNİK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ LEİSHMANİA PARAZİTLERİNDEN LİPOFOSFOGLİKAN (LPG) SAFLAŞTIRILMASI VE AŞI MODELİ OLARAK LPG-POLİMER KONJUGATININ ANTİLEİSHMANİAL ETKİNLİĞİNİN IN VITRO
Yrd. Doç. Dr. İlyas Yolbaş Çocuk Sağlığı ve Hastalıkları ABD
 Yrd. Doç. Dr. İlyas Yolbaş Çocuk Sağlığı ve Hastalıkları ABD KOMPLEMAN SİSTEMİ Kompleman sistem, (Compleman system) veya tamamlayıcı sistem, bir canlıdan patojenlerin temizlenmesine yardım eden biyokimyasal
Yrd. Doç. Dr. İlyas Yolbaş Çocuk Sağlığı ve Hastalıkları ABD KOMPLEMAN SİSTEMİ Kompleman sistem, (Compleman system) veya tamamlayıcı sistem, bir canlıdan patojenlerin temizlenmesine yardım eden biyokimyasal
BİRİNCİ BASAMAKTA PRİMER İMMÜN YETMEZLİK
 1 LERDE LABORATUVAR İPUÇLARI GENEL TARAMA TESTLERİ Tam kan sayımı Periferik yayma İmmünglobulin düzeyleri (IgG, A, M, E) İzohemaglutinin titresi (Anti A, Anti B titresi) Aşıya karşı antikor yanıtı (Hepatit
1 LERDE LABORATUVAR İPUÇLARI GENEL TARAMA TESTLERİ Tam kan sayımı Periferik yayma İmmünglobulin düzeyleri (IgG, A, M, E) İzohemaglutinin titresi (Anti A, Anti B titresi) Aşıya karşı antikor yanıtı (Hepatit
HUMORAL İMMUN YANIT 1
 HUMORAL İMMUN YANIT 1 Antijen B lenfosit... HUMORAL İMMUN YANIT Antikor üretimi 2 Antijenini işlenmesi ve sunulması Yardımcı T-lenfosit aktivasyonu Yardımcı T hücre- B hücre ilişkisi B hücre aktivasyonu
HUMORAL İMMUN YANIT 1 Antijen B lenfosit... HUMORAL İMMUN YANIT Antikor üretimi 2 Antijenini işlenmesi ve sunulması Yardımcı T-lenfosit aktivasyonu Yardımcı T hücre- B hücre ilişkisi B hücre aktivasyonu
Leishmania nın Türkiye deki Epidemiyolojisi ve Tanısı
 Leishmania nın Türkiye deki Epidemiyolojisi ve Tanısı Prof. Dr. Seray TÖZ Ege Üniversitesi Tıp Fakültesi Parazitoloji Anabilim Dalı, Bornova, Izmir seray.ozensoy.toz@ege.edu.tr İnsanlarda Leishmaniasis
Leishmania nın Türkiye deki Epidemiyolojisi ve Tanısı Prof. Dr. Seray TÖZ Ege Üniversitesi Tıp Fakültesi Parazitoloji Anabilim Dalı, Bornova, Izmir seray.ozensoy.toz@ege.edu.tr İnsanlarda Leishmaniasis
DÖNEM 2- I. DERS KURULU AMAÇ VE HEDEFLERİ
 DÖNEM 2- I. DERS KURULU AMAÇ VE HEDEFLERİ Kan, kalp, dolaşım ve solunum sistemine ait normal yapı ve fonksiyonların öğrenilmesi 1. Kanın bileşenlerini, fiziksel ve fonksiyonel özelliklerini sayar, plazmanın
DÖNEM 2- I. DERS KURULU AMAÇ VE HEDEFLERİ Kan, kalp, dolaşım ve solunum sistemine ait normal yapı ve fonksiyonların öğrenilmesi 1. Kanın bileşenlerini, fiziksel ve fonksiyonel özelliklerini sayar, plazmanın
Deri Layşmanyazisi. Prof. Dr. Mehmet HARMAN Dicle Üniversitesi Tıp Fakültesi Dermatoloji Anabilim Dalı
 Deri Layşmanyazisi Prof. Dr. Mehmet HARMAN Dicle Üniversitesi Tıp Fakültesi Dermatoloji Anabilim Dalı Amaç Hastalığın tanısını koyabilmek Uygun tedaviyi yapabilmek Koruyucu yöntemleri sayabilmek İçerik
Deri Layşmanyazisi Prof. Dr. Mehmet HARMAN Dicle Üniversitesi Tıp Fakültesi Dermatoloji Anabilim Dalı Amaç Hastalığın tanısını koyabilmek Uygun tedaviyi yapabilmek Koruyucu yöntemleri sayabilmek İçerik
Bağ doku. Mezodermden köken alır. En Yaygın bulunan dokudur ( Epitel, Kas, Kemik sinir)
") Bağ doku Mezodermden köken alır En Yaygın bulunan dokudur ( Epitel, Kas, Kemik sinir) Bağ dokunun Fonksiyonları Diğer organ ve dokuların Fonksiyonal ve yapısal desteğini sağlar. kan damarları aracılığı
Bağ doku Mezodermden köken alır En Yaygın bulunan dokudur ( Epitel, Kas, Kemik sinir) Bağ dokunun Fonksiyonları Diğer organ ve dokuların Fonksiyonal ve yapısal desteğini sağlar. kan damarları aracılığı
2003 ÖSS BİYOLOJİ SORULARI VE CEVAPLARI
 2003 ÖSS BİYOLOJİ SORULARI VE CEVAPLARI 1. Bir hücrede oksijenli solunum, protein sentezi, fotosentez olaylarının tümünün gerçekleşebilmesi için, bu hücrede; I. ribozom, II. kloroplast, III. mitokondri,
2003 ÖSS BİYOLOJİ SORULARI VE CEVAPLARI 1. Bir hücrede oksijenli solunum, protein sentezi, fotosentez olaylarının tümünün gerçekleşebilmesi için, bu hücrede; I. ribozom, II. kloroplast, III. mitokondri,
Yrd. Doç. Dr. Tuba ŞANLI
 Yrd. Doç. Dr. Tuba ŞANLI Genel olarak gözle net olarak görülemeyecek kadar küçük canlıları inceleyen ve onları konu olarak ele alan bilim dalıdır. Gözle ayırt edilemeyen canlılar; Virüsler, bakteriler,
Yrd. Doç. Dr. Tuba ŞANLI Genel olarak gözle net olarak görülemeyecek kadar küçük canlıları inceleyen ve onları konu olarak ele alan bilim dalıdır. Gözle ayırt edilemeyen canlılar; Virüsler, bakteriler,
Mikroorganizmalar gözle görülmezler, bu yüzden mikroskopla incelenirler.
 10. Sınıf Enfeksiyondan Korunma 2.Hafta ( 22-26 / 09 / 2014 ) ENFEKSİYON ETKENLERİNİN SINIFLANDIRILMASI 1.) BAKTERİLER 2.) VİRÜSLER Slayt No : 2 Mikroorganizmaların Sınıflandırılması ; a.) Sayısal Yöntem,
10. Sınıf Enfeksiyondan Korunma 2.Hafta ( 22-26 / 09 / 2014 ) ENFEKSİYON ETKENLERİNİN SINIFLANDIRILMASI 1.) BAKTERİLER 2.) VİRÜSLER Slayt No : 2 Mikroorganizmaların Sınıflandırılması ; a.) Sayısal Yöntem,
T.C. MUSTAFA KEMAL ÜNİVERSİTESİ TAYFUR ATA SÖKMEN TIP FAKÜLTESİ EĞİTİM & ÖĞRETİM YILI DÖNEM II
 T.C. MUSTAFA KEMAL ÜNİVERSİTESİ TAYFUR ATA SÖKMEN TIP FAKÜLTESİ 2017 2018 EĞİTİM & ÖĞRETİM YILI DÖNEM II VI. DERS KURULU (4 HAFTA) 1901206HASTALIKLARIN BİYOLOJİK TEMELİ DERS KURULU-I DEKAN DEKAN YRD. BAŞKORDİNATÖR
T.C. MUSTAFA KEMAL ÜNİVERSİTESİ TAYFUR ATA SÖKMEN TIP FAKÜLTESİ 2017 2018 EĞİTİM & ÖĞRETİM YILI DÖNEM II VI. DERS KURULU (4 HAFTA) 1901206HASTALIKLARIN BİYOLOJİK TEMELİ DERS KURULU-I DEKAN DEKAN YRD. BAŞKORDİNATÖR
BELKİDE BİYOLOJİNİN EN TEMEL KONUSU EN ZEVKLİ KONUSUNA BAŞLAYALIM ARKADAŞLAR!!!
 DERS : BİYOLOJİ KONU: HÜCRE BELKİDE BİYOLOJİNİN EN TEMEL KONUSU EN ZEVKLİ KONUSUNA BAŞLAYALIM ARKADAŞLAR!!! Canlıların canlılık özelliği gösteren en küçük yapı birimidir.( Virüsler hariç) Şekil: Bir hayvan
DERS : BİYOLOJİ KONU: HÜCRE BELKİDE BİYOLOJİNİN EN TEMEL KONUSU EN ZEVKLİ KONUSUNA BAŞLAYALIM ARKADAŞLAR!!! Canlıların canlılık özelliği gösteren en küçük yapı birimidir.( Virüsler hariç) Şekil: Bir hayvan
*Canlıların canlılık özelliği gösteren en küçük yapı birimine hücre denir.
 Fen ve Teknoloji 1. Ünite Özeti Hücre Canlılarda Üreme, Büyüme ve Gelişme. *Canlıların canlılık özelliği gösteren en küçük yapı birimine hücre denir. *Hücrenin temel kısımları: hücre zarı, sitoplâzma ve
Fen ve Teknoloji 1. Ünite Özeti Hücre Canlılarda Üreme, Büyüme ve Gelişme. *Canlıların canlılık özelliği gösteren en küçük yapı birimine hücre denir. *Hücrenin temel kısımları: hücre zarı, sitoplâzma ve
Bütün hücrelerin olmazsa olmazları. Plazma zarı Yarı-sıvı sitosol Kromozom Ribozom
 Bütün hücrelerin olmazsa olmazları Plazma zarı Yarı-sıvı sitosol Kromozom Ribozom Hücre Ökaryotik hücre Nukleus var Zarla çevrili organeller var Genellikle prokaryotik hücreye oranla daha büyüktür. Ökaryotlardaki
Bütün hücrelerin olmazsa olmazları Plazma zarı Yarı-sıvı sitosol Kromozom Ribozom Hücre Ökaryotik hücre Nukleus var Zarla çevrili organeller var Genellikle prokaryotik hücreye oranla daha büyüktür. Ökaryotlardaki
HEREDİTER SFEROSİTOZ. Mayıs 14
 HEREDİTER SFEROSİTOZ İNT.DR.DİDAR ŞENOCAK Giriş Herediter sferositoz (HS), hücre zarı proteinlerinin kalıtsal hasarı nedeniyle, eritrositlerin morfolojik olarak bikonkav ve santral solukluğu olan disk
HEREDİTER SFEROSİTOZ İNT.DR.DİDAR ŞENOCAK Giriş Herediter sferositoz (HS), hücre zarı proteinlerinin kalıtsal hasarı nedeniyle, eritrositlerin morfolojik olarak bikonkav ve santral solukluğu olan disk
D Vitaminin Relaps Brucelloz üzerine Etkisi. Yrd.Doç.Dr. Turhan Togan Başkent Üniversitesi Enfeksiyon Hastalıkları ve Klinik Mikrobiyoloji
 D Vitaminin Relaps Brucelloz üzerine Etkisi Yrd.Doç.Dr. Turhan Togan Başkent Üniversitesi Enfeksiyon Hastalıkları ve Klinik Mikrobiyoloji Bruselloz Brucella cinsi bakteriler tarafından primer olarak otçul
D Vitaminin Relaps Brucelloz üzerine Etkisi Yrd.Doç.Dr. Turhan Togan Başkent Üniversitesi Enfeksiyon Hastalıkları ve Klinik Mikrobiyoloji Bruselloz Brucella cinsi bakteriler tarafından primer olarak otçul
Steril Hücre Kültürü Tekniği h"ps://www.youtube.com/watch?v=ugcgo42vnqi
 Kontaminasyon Steril Hücre Kültürü Tekniği h"ps://www.youtube.com/watch?v=ugcgo42vnqi Kontaminasyon Kontaminasyon hücre kültürü laboratuvarlarında en çok karşılaşılan problemlerden birisidir. Hücrelerin
Kontaminasyon Steril Hücre Kültürü Tekniği h"ps://www.youtube.com/watch?v=ugcgo42vnqi Kontaminasyon Kontaminasyon hücre kültürü laboratuvarlarında en çok karşılaşılan problemlerden birisidir. Hücrelerin
CANLILARIN ORTAK ÖZELLİKLERİ
 1 CANLILARIN ORTAK ÖZELLİKLERİ 1.Hücresel yapıdan oluşur 2.Beslenir 3.Solunum yapar 4.Boşaltım yapar 5.Canlılar hareket eder 6.Çevresel uyarılara tepki gösterir 7.Büyür ve gelişir (Organizasyon) 8.Üreme
1 CANLILARIN ORTAK ÖZELLİKLERİ 1.Hücresel yapıdan oluşur 2.Beslenir 3.Solunum yapar 4.Boşaltım yapar 5.Canlılar hareket eder 6.Çevresel uyarılara tepki gösterir 7.Büyür ve gelişir (Organizasyon) 8.Üreme
TIBBİ MİKROBİYOLOJİ ANABİLİM DALI MEZUNİYET SONRASI (UZMANLIK) EĞİTİMİ DERS MÜFREDATI
 EĞİTİMİ DERS MÜFREDATI") TIBBİ MİKROBİYOLOJİ ANABİLİM DALI MEZUNİYET SONRASI (UZMANLIK) EĞİTİMİ DERS MÜFREDATI DERS KODU MİK 0001 MİK 0002 MİK 0003 MİK 0004 MİK 0005 MİK 0006 MİK 0007 MİK 1000 MİK 4000 MİK 5000 KONU ADI Vaka Değerlendirme
TIBBİ MİKROBİYOLOJİ ANABİLİM DALI MEZUNİYET SONRASI (UZMANLIK) EĞİTİMİ DERS MÜFREDATI DERS KODU MİK 0001 MİK 0002 MİK 0003 MİK 0004 MİK 0005 MİK 0006 MİK 0007 MİK 1000 MİK 4000 MİK 5000 KONU ADI Vaka Değerlendirme
Tüm yaşayan organizmalar suya ihtiyaç duyarlar Çoğu hücre suyla çevrilidir ve hücrelerin yaklaşık %70 95 kadarı sudan oluşur. Yerküre içerdiği su ile
 Su Kimyası Tüm yaşayan organizmalar suya ihtiyaç duyarlar Çoğu hücre suyla çevrilidir ve hücrelerin yaklaşık %70 95 kadarı sudan oluşur. Yerküre içerdiği su ile canlılık için gerekli ortamı sunar. Canlıların
Su Kimyası Tüm yaşayan organizmalar suya ihtiyaç duyarlar Çoğu hücre suyla çevrilidir ve hücrelerin yaklaşık %70 95 kadarı sudan oluşur. Yerküre içerdiği su ile canlılık için gerekli ortamı sunar. Canlıların
Prof.Dr.Kemal NAS Dicle Üniversitesi Tıp Fakültesi Fizik Tedavi ve Rehabilitasyon AD, Romatoloji BD
 Prof.Dr.Kemal NAS Dicle Üniversitesi Tıp Fakültesi Fizik Tedavi ve Rehabilitasyon AD, Romatoloji BD Kronik enflamatuar hastalıklar, konak doku ve immun hücreleri arasındaki karmaşık etkileşimlerinden
Prof.Dr.Kemal NAS Dicle Üniversitesi Tıp Fakültesi Fizik Tedavi ve Rehabilitasyon AD, Romatoloji BD Kronik enflamatuar hastalıklar, konak doku ve immun hücreleri arasındaki karmaşık etkileşimlerinden
Bilimsel Projeler. Projenin amacı: Öğrencileri okul tuvaletlerinde mikroorganizmaların bulaşarak hasta olmalarını engellemek,
 Bilimsel Projeler PROJENİN ADI: Mikroorganizmaları Işınlayalım Öğrenciler: Rumeysa ŞAHİN Okul: Amasya Bilim ve Sanat Merkezi - 2012 Danışman: Hidayet TERECİ www.fencebilim.com Projenin amacı: Öğrencileri
Bilimsel Projeler PROJENİN ADI: Mikroorganizmaları Işınlayalım Öğrenciler: Rumeysa ŞAHİN Okul: Amasya Bilim ve Sanat Merkezi - 2012 Danışman: Hidayet TERECİ www.fencebilim.com Projenin amacı: Öğrencileri
I. DÖNEM - 2. DERS KURULU ( )
") Açıklamalar: I. DÖNEM - 2. DERS KURULU (2014-2015) Kısaltmalar: DK: Ders kurulu, IHU: İyi hekimlik uygulamaları, Mİng: Akademik/Medikal İngilizce, TDE: Türk Dili ve Edebiyatı, Bilgisayar Okur yazarlığı:
Açıklamalar: I. DÖNEM - 2. DERS KURULU (2014-2015) Kısaltmalar: DK: Ders kurulu, IHU: İyi hekimlik uygulamaları, Mİng: Akademik/Medikal İngilizce, TDE: Türk Dili ve Edebiyatı, Bilgisayar Okur yazarlığı:
Mikrobiyal Gelişim. Jenerasyon süresi. Bakterilerde üreme eğrisi. Örneğin; (optimum koşullar altında) 10/5/2015
 10/5/2015") Mikrobiyal Gelişim Tek hücreli organizmalarda sayı artışı Bakterilerde en çok görülen üreme şekli ikiye bölünmedir (mikroorganizma sayısı) Çok hücreli organizmalarda kütle artışı Genelde funguslarda görülen
Mikrobiyal Gelişim Tek hücreli organizmalarda sayı artışı Bakterilerde en çok görülen üreme şekli ikiye bölünmedir (mikroorganizma sayısı) Çok hücreli organizmalarda kütle artışı Genelde funguslarda görülen
BİYOLOJİK MOLEKÜLLERDEKİ
 BİYOLOJİK MOLEKÜLLERDEKİ KİMYASALBAĞLAR BAĞLAR KİMYASAL VE HÜCRESEL REAKSİYONLAR Yrd. Doç.Dr. Funda BULMUŞ Atomun Yapısı Maddenin en küçük yapı taşı olan atom elektron, proton ve nötrondan oluşmuştur.
BİYOLOJİK MOLEKÜLLERDEKİ KİMYASALBAĞLAR BAĞLAR KİMYASAL VE HÜCRESEL REAKSİYONLAR Yrd. Doç.Dr. Funda BULMUŞ Atomun Yapısı Maddenin en küçük yapı taşı olan atom elektron, proton ve nötrondan oluşmuştur.
TIBBĠ BĠLĠMLERE GĠRĠġ DĠLĠMĠ MĠKROBĠYOLOJĠ ANABĠLĠM DALI
 TIBBĠ BĠLĠMLERE GĠRĠġ DĠLĠMĠ MĠKROBĠYOLOJĠ ANABĠLĠM DALI ÖĞRETĠM ÜYESĠ : Prof. Dr. O. ġadi Yenen Ders: VĠROLOJĠYE GĠRĠġ, TARĠHÇE ve EVRĠM 1. Virusların tanımlanması ve rolüne ilişkin önemli tarihsel gelişmelerin
TIBBĠ BĠLĠMLERE GĠRĠġ DĠLĠMĠ MĠKROBĠYOLOJĠ ANABĠLĠM DALI ÖĞRETĠM ÜYESĠ : Prof. Dr. O. ġadi Yenen Ders: VĠROLOJĠYE GĠRĠġ, TARĠHÇE ve EVRĠM 1. Virusların tanımlanması ve rolüne ilişkin önemli tarihsel gelişmelerin