ANAPLASTİK BEYİN TÜMÖRLERİNİN TANI VE PROGNOZUNDA TÜMÖR SÜPRESSÖR GENLERİN VE ONKOGENLERİN ÖNEMİ
|
|
|
- Erol Poyraz
- 5 yıl önce
- İzleme sayısı:
Transkript
1 T.C. EGE ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ ANAPLASTİK BEYİN TÜMÖRLERİNİN TANI VE PROGNOZUNDA TÜMÖR SÜPRESSÖR GENLERİN VE ONKOGENLERİN ÖNEMİ Tıbbi Biyoloji Anabilim Dalı Programı Doktora Tezi Araştırma Görevlisi Çığır BİRAY AVCI DANIŞMAN Doç. Dr. Cumhur GÜNDÜZ İZMİR 2009
2 T.C. EGE ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ ANAPLASTİK BEYİN TÜMÖRLERİNİN TANI VE PROGNOZUNDA TÜMÖR SÜPRESSÖR GENLERİN VE ONKOGENLERİN ÖNEMİ Tıbbi Biyoloji Anabilim Dalı Programı Doktora Tezi Araştırma Görevlisi Çığır BİRAY AVCI DANIŞMAN Doç. Dr. Cumhur GÜNDÜZ İZMİR 2009
3
4 DEĞERLENDİRME KURULU ÜYELERİ (Adı Soyadı) (İmza) Başkan : Doç. Dr. Cumhur GÜNDÜZ (Danışman)... Üye : Prof. Dr. Nejat TOPÇUOĞLU Üye : Prof. Dr. Meral SAKIZLI... Üye : Prof. Dr. Nezih OKTAR... Üye : Doç. Dr. Özgür ÇOĞULU. Doktora Tezinin kabul edildiği tarih:.../.../.2009
5 ÖNSÖZ Yüksek lisans ve doktora eğitimim süresince değerli görüşlerini, bilgilerini ve desteğini hiçbir zaman esirgemeyen yanında zevkle çalıştığım, babacan tavırları ve hoşgörüsüyle her zaman yanımda hissettiğim, Anabilim Dalı Başkanımız Sayın Prof.Dr. Nejat TOPÇUOĞLU na, Gerek tıbbi gerekse sosyal alanlarda eşsiz deneyimlerini benimle paylaşan, yol göstericiliği, sabrı, yaratıcılığı, içtenliği ve desteğini her an hissettiğim danışmanım Sayın Doç. Dr. Cumhur GÜNDÜZ e, Samimi tavırları ile her zaman desteğini hissedip rahatladığım, Anabilim Dalımızın değerli öğretim üyesi Yard. Doç. Dr. Sayın Zuhal EROĞLU na, Anabilim Dalımızın öğretim üyelerinden Yard. Doç. Dr. Sayın Buket KOSOVA ya, Nöroşirürji bilimi ile tanışmamı ve nöroonkolojiye yönlenmemi sağlayan, tezimin proje haline getirilmesinden, tümör olgu gruplarının toplanmasına kadar her yönüyle beni teşvik edip, destekleyen değerli Sayın hocam Prof. Dr. Nezih OKTAR a, Tezimin proje haline getirilmesinde yol göstericiliği ve bilimsel katkısının yanında, tümör olgu gruplarının çoğunluğunun opere edilmesini sağlayan, Sayın abim değerli Prof. Dr. Tayfun DALBASTI ya,
6 Patoloji ye gönderdiğim örneklerin tamamının tanı ve derece takibini yapan, her sorumu anında yanıtlayan Sayın hocam Prof. Dr. Taner AKALIN a, Tezimin proje haline getirilmesi ve çalışmalarımın moleküler genetik aşamalarının yönlendirilmesinde katkıda bulunan değerli abim, Sayın hocam Doç. Dr. Özgür ÇOĞULU ya, İçten dostluklarını her an hissettiğim, birlikte neredeyse Dünyayı gezdiğimiz, tezimin proje haline getirilmesinden, laboratuvar çalışmalarının birebir takibine ve yazım aşamalarının her anında destek ve yürek veren dostlarım ve çalışma arkadaşlarım Ar. Gör. Sayın Sunde YILMAZ, Doktora öğrencisi Sayın Z. Özlem DOĞAN a, Koşulsuz yardımları ve dostluğu için değerli arkadaşım Ar. Gör. Sayın Yavuz DODURGA ya, Bilimsel olarak ayrıldığım ama gönül bağlarımızın Katar a kadar uzandığı, bitmeyen enerjisi ve eşsiz dostluğunu her an hissettiren canım kardeşim, Yüksek BiyoMühendis Sinem Numanoğlu GÜNDÜZ e, Asistanlığım süresince destekleyici ve yüreklendirici tavırları, destek ve yardımları, birlikte paylaştığımız tüm güzellikler için çalışma arkadaşlarım Uzman Dr. Sayın Nur SELVİ ye, Ar. Gör. Sayın Burçin TEZCANLI ya, Ar. Gör. Sayın Vildan BOZOK ÇETINTAŞ a, Ar. Gör. Sayın Aslı TETİK e, Ar. Gör. Sayın Duygu AYGÜNEŞ e, Yeni yüksek lisans eğitimine başlamasına rağmen, sempatik tavırları ve çalışkanlığıyla sevgimi kazanan Sayın Müsteyde YÜCEBAŞ a, II
7 İstediğim her an tezimin malzemelerini temin eden ve desteğiyle beni hep yüreklendiren canım arkadaşım Sayın Bircan İSCA ya, Anabilim Dalımızın çok değerli ve çalışkan sekreterleri, ameliyathaneden gelen her telefonu bildiren ve tümörlerin elime ulaşmasını birebir takip eden canım ablam Sayın Gönül SAKLICA ya ve biricik Sayın Anıl KARAKAŞ a, Değerli personelimiz Sayın Salih TOROĞLU ya, Tez çalışmamın projesine maddi destek sağlayan TÜBİTAK a, İyi ve kötü, her anımda yanımda olan ve bana sonsuz destek veren, en kötü anımda beni mutlu edebilen, saygı ve sevgisini hiçbir zaman esirgemeyen canım eşim Makina Mühendisi Sayın Tolga AVCI ya, Mesafelere rağmen, her an sıcacık yüreğiyle hep yanımda olan, desteğini hiçbir zaman esirgemeyen, en zor anlarımda beni yüreklendirerek aydınlığa çıkaran, ablası olmaktan her zaman gurur duyduğum biricik kardeşim Çağıl BİRAY a, Tezimin son aşamalarında varlığını öğrenmemle hayatımı değiştiren, tezi kaleme döktüğüm bu evrelerde dahi beni hiç üzmeyen, yalnızca karnımda hafif hareketlerle varlığını hissettiren 21 haftalık eşsiz BEBEĞİM e, Hayatımın her saniyesinde sorgusuz ve koşulsuz desteklerini hissettiğim, beni ben yapan öğelerin mimarları, canım ANNE ve BABAMA, SONSUZ TEŞEKKÜRLER.. İzmir, 10 Haziran 2009 Araştırma Gör. Çığır Biray AVCI III
8 BÖLÜM I İÇİNDEKİLER 1.1 GİRİŞ GENEL BİLGİLER Beyin (Encephalon) Beyin Tümörleri Epidemiyoloji Olguların Yaş ve Cinsiyeti Beyin Tümör Tiplerinin Histolojisi ve Moleküler Biyolojisi Sinir Sistemi Tümörlerinin Sınıflandırılması Dünya Sağlık Örgütü (WHO) Derecelendirmesi Gliomlar Astrositomlar Oligodendrogliomlar Ependimomlar Meningiomlar Medulloblastomlar Ganglioglioma Schwannomlar Kordomalar Nörofibromatozis Nörofibromatozis Tip Yeni Tanımlanan Tümörler Atipik Koroid Pleksus Papillomu Ekstraventriküler Nörositoma Papiller Glionöronal Tümör Ventrikülün Rozet Oluşturan Glionöronal Tümörü Pineal Bölgenin Papiller Tümörü Pütisitom Adenohipofizin İğsi Hücreli Onkositomu Beyin Tümörlerinin Ailesel İlişkileri Beyin Tümörlerinin Ailesel İlişkileri Ailesel Yığılma Metabolik Yatkınlık ve Polimorfizmler Tümör Süpresör Genler p RB1 (Retinoblastoma) PTEN (Fosfataz ve tensin homoloğu, MMAC 1) MGMT (O 6-metilguanin-DNA metiltransferaz) RUNX3 (Runt ile ilişkili transkripsiyon faktörü 3) DMBT1 (Malin Beyin Tümörlerinde Silinmiş 1 Geni) PIKE (Fosfotidilinositol 3-kinaz arttırıcı, CENTG1) 44 IV
9 Onkogenler EGFR (Epidermal Büyüme Faktörü Reseptörü) PIK3CA (Fosfoinosit 3 Kinaz, Katalitik, Alfa Polipeptit) MDM2 [MDM2 p53 Binding Protein Homolog (Mouse)] OLIG2 (Oligodendrosit Transkripsiyon Faktörü 2) GSTT1 (Glutatyon S-Transferaz Teta 1) COX 2 (Siklooksigenaz 2) htert (İnsan Telomeraz Revers Transkriptazı) Epigenetik Epigenetik Yığılma Metilasyon Metilasyona Özgü PCR Beyin Tümörü Modelleri Gen Ekspresyonu ve Gen Kopya Sayısı Eksklüzyon Boyaları Tripan Mavisi Boyası Canlılık Testi 76 BÖLÜM II 2.1. GEREÇ VE YÖNTEM Hücre Kültürü ve Moleküler Çalışmalarda Kullanılan Cihazlar ve Kimyasallar Olgu ve Hücre Hatları Tümör Derece ve Evresinin Belirlenmesi Tümör Hücre Kültürü Hücre Yedeklenmesi Tümör Hücrelerinin ve Hücre Hatlarının Hücrelerinin Pasajlanması Tümör Hücrelerinin ve Hücre hatlarının Canlılığının Belirlenmesi Total RNA İzolasyonu Revers Transkriptaz RT-PCR cdna Sentezi Real-time Online Revers Transkriptaz RT-PCR ve Taqman Protokolü Tümör Süpresör ve Onkogen Gen Ekspresyon Analizlerinde Kullanılan Primer ve Problar Ekspresyonun Değerlendirilmesi Gen Kopya Sayısı DNA İzolasyonu Gen kopya sayısının Kantitatif Analizleri Tümör Süpresör Genlerin Hipermetilasyonun Kantitasyon Analizleri Bisülfit Uygulaması DNA metilasyonu Protokolü Genlerin Kantitasyon Değerlerinin İstatistiksel Analizleri 109 V
10 BÖLÜM III 3.1. BULGULAR Olguların Belirlenmesi ve Tümör dokusu alımı Tümör derece ve evresinin belirlenmesi Gen Ekspresyon Analizleri Onkogenler PIK3CA MDM GSTT EGFR OLIG COX htert Tümör süpresör genler Gen Kopya Sayısı Değişimleri Onkogen Kopya Sayısı Değişimleri Tümör Süpresör gen Kopya Sayısı Değişimleri Tümör Süpresör Genlerin Hipermetilasyonun Kantitasyon Analizleri Onkogen, Tümör Süpresör Gen Ekspresyonları, Kopya Sayıları ve Tümör Süpresör Gen Hipermetilasyonunun Ayırt Ediciliği Onkogen ve Tümör Süpresör Genler ile Sağkalım İlişkileri 215 BÖLÜM IV 4.1. TARTIŞMA 228 BÖLÜM V 5.1. SONUÇ ve ÖNERİLER 259 BÖLÜM VI 6.1. ÖZET ABSTRACT 267 BÖLÜM VII 7.1. YARARLANILAN KAYNAKLAR 268 ÖZGEÇMİŞ 294 VI
11 Tablo dizini Tablo 1: Kullanılan cihazlar 78 Tablo 2: Kullanılan kimyasal maddeler 79 Tablo 3: Komplementer DNA analizinin ilk basamağı 89 Tablo 4: cdna analizinin ikinci basamağı 89 Tablo 5: Tümör süpresörler için belirlenen İlgili Primer ve problar 90 Tablo 6: Onkogenler için belirlenen İlgili Primer ve problar 91 Tablo 7: Primer prob dizaynı ile ilgili Açıklama 97 Tablo 8: Metilasyon için gerekli primer ve probların dizaynı ile ilgili açıklama 103 Tablo 9: Beyin tümörlü olgu gruplarının demografik özellikleri 111 Tablo 10: Olguların klinik özellikleri 112 Tablo 11: Olguların tanı gruplarındaki cinsiyet dağılımı 114 Tablo 12: Tümör olgu gruplarının yaş dağılımı 115 Tablo 13: PIK3CA gen ekspresyonunun rölatif oranlarının ortalaması ve %95 Güven aralığı 117 Tablo 14: Pozitif ve negatif kontrol hücre hatlarındaki PIK3CA gen ekspresyonu 119 Tablo 15: MDM2 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 Güven aralığı 120 Tablo 16: Pozitif ve negatif kontrol hücre hatlarındaki MDM2 gen ekspresyonu 120 Tablo 17: GSTT1 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 Güven aralığı 122 Tablo 18: GSTT1 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 Güven aralığı 123 Tablo 19: EGFR gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 Güven aralığı 125 Tablo 20: EGFR gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 Güven aralığı 125 Tablo 21: OLIG2 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 Güven aralığı 127 Tablo 22: OLIG2 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 Güven aralığı 128 Tablo 23: COX-2 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 Güven aralığı 130 Tablo 24: COX-2 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 Güven aralığı 130 Tablo 25: htert gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 Güven aralığı 133 Tablo 26: htert gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 Güven aralığı 133 Tablo 27: Anaplastik Astrositom grubunda onkogenlerin karşılıklı ilişkileri 136 Tablo 28: Anaplastik Oligodendrogliom grubunda onkogenlerin birbiri ile etkileşimleri Tablo 29: Diffüz Astrositom grubunda onkogenlerin birbiri ile etkileşimleri 137 Tablo 30: Glioblastom grubunda onkogenlerin birbiri ile etkileşimi 139 VII
12 Tablo 31: Meningiom grubunda onkogenlerin birbiri ile etkileşimi 140 Tablo 32: Tümör derecelerine göre onkogen ekspresyon değişimleri 141 Tablo 33: Tümör süpresör gen ekspresyonlarının rölatif oranlarının tümör grubu ortalaması ve %95 Güven aralığı 143 Tablo 34: Tümör süpresör gen ekspresyonlarının rölatif oranlarının hücre hatlarındaki ortalaması ve %95 Güven aralığı 157 Tablo 35: Anaplastik Oligodendrogliom grubunda onkogenler ile tümör süpresör genlerin karşılıklı etkileşimleri 158 Tablo 36: Diffüz astrositom grubunda onkogenler ile tümör süpresör genlerin karşılıklı etkileşimleri 159 Tablo 37: Glioblastom grubunda onkogenler ile tümör süpresör genlerin karşılıklı etkileşimleri 160 Tablo 38: Meningiom grubunda onkogenler ile tümör süpresör genlerin karşılıklı etkileşimleri 161 Tablo 39: Oligodendrogliom grubunda onkogenler ile tümör süpresör genlerin karşılıklı etkileşimleri 162 Tablo 40: Schwannom grubunda onkogenler ile tümör süpresör genlerin karşılıklı etkileşimleri 163 Tablo 41: Tümör süpresör genlerin tümör derecelerine göre sınıflandırılmaları 164 Tablo 42: Tümör grupları arasında onkogen kopya sayısı dağılımı 166 Tablo 43: Negatif kontrol hücre hatları ile glioblastom hücre hattı karşılaştırılması 170 Tablo 44: Tümör grupları arasında tümör süpresör gen kopya sayısı dağılımı 178 Tablo 45: Negatif ve pozitif kontrol grupları arasında tümör süpresör gen kopya sayısı dağılımı 182 Tablo 46: Tümör olgularının tümör süpresör gen promotor bölgelerinin metile/unmetile ve metile/mix rölatif oranları 194 Tablo 47: Negatif ve pozitif kontrol gruplarının tümör süpresör gen promotor bölgelerinin metile/unmetile ve metile/mix rölatif oranları 199 Tablo 48: RB1 M/UM metilasyon skalasına göre dağılım 210 Tablo 49: PTEN M/UM metilasyon skalasına göre dağılım 211 Tablo 50: RUNX3 M/UM metilasyon skalasına göre dağılım 212 Tablo 51: MGMT M/UM metilasyon skalasına göre dağılım 213 Tablo 52: Astrositk tümörlerde tümör süpresör gen ve onkogen ekspresyon, gen kopya sayı ve gen promotör bölge metilasyon değerlerinin olguların kliniği ile korelasyonu 214 Tablo 53: Oligodendroglial tümörlerde tümör süpresör gen ve onkogen ekspresyon, gen kopya sayı ve gen promotör bölge metilasyon değerlerinin olguların kliniği ile korelasyonu 215 Tablo 54: Onkogen ekspresyon pozitifliğine göre tahmini sağkalım ile ilişkisi 216 Tablo 55: Tümör süpresör gen kopya sayısı aberasyonuna göre tahmini sağkalım ile ilişkisi 220 Tablo 56: Onkogen ekspresyon pozitifliğinin tahmini sağkalım ile ilişkisi 225 VIII
13 Tablo 57: Tümör süpresör gen ekspresyon pozitifliğinin tahmini sağkalım ile ilişkisi 227 IX
14 Şekil Dizini Şekil 1: Beynin üstten ve alttan görünüşü 4 Şekil 2: Glioma patogenezi 18 Şekil 3: Neubauer hemositometresi 77 Şekil 4: Beyin tümörlü olgunun hücre kültürünün inverted mikroskoptaki görüntüsü 81 Şekil 5: Kültüre edilen U-87 MG hücre hattının inverted mikroskop görüntüsü 82 Şekil 6: Çalışma kapsamında uygulanan yöntem 83 Şekil 7: Eksklüzyon boya prensibi 86 Şekil 8: (a) Hedef Gen, (b) housekeeping Gen Geçiş Noktası 95 Şekil 9: Metilasyon için gerekli primer ve probların dizaynı 101 X
15 Grafik Dizini Grafik 1: tümör olgu gruplarının cinsiyete göre dağılımı 111 Grafik 2: olguların tanılarına göre dağılımı 113 Grafik 3: olguların tanı gruplarına göre cinsiyet dağılımlarının yüzde değişimleri 115 Grafik 4:olguların tanılarına göre yaş ortalamaları 116 Grafik 5: pık3ca gen ekspresyonu 118 Grafik 6: pozitif ve negatif kontrol hücre hatlarındaki pık3ca gen ekspresyonu 119 Grafik 7: mdm2 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 121 Grafik 8: mdm2 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 121 Grafik 9: gstt1 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 123 Grafik 10: gstt1 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 124 Grafik 11: egfr gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 126 Grafik 12: egfr gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 126 Grafik 13: olıg2 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 128 Grafik 14: olıg2 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 129 Grafik 15: cox-2 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 131 Grafik 16: cox-2 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 131 Grafik 17: htert gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 134 Grafik 18: htert gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 134 Grafik 19: dmbt1 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 148 Grafik 20: dmbt1 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 148 Grafik 21: mgmt gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 149 Grafik 22: mgmt gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 149 Grafik 23: pıke gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 150 Grafik 24: pıke gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 150 Grafik 25: pten gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 151 XI
16 Grafik 26: pten gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 151 Grafik 27: rb1 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 152 Grafik 28: rb1 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 152 Grafik 29: runx3 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 153 Grafik 30: runx3 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 153 Grafik 31: p53 gen ekspresyonunun rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 154 Grafik 32: p53 gen ekspresyonunun rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 154 Grafik 33: pık3ca gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 171 Grafik 34: pık3ca gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 171 Grafik 35: mdm2 gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 172 Grafik 36: mdm2 gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 172 Grafik 37: gstt1 gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 173 Grafik 38: gstt1 gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 173 Grafik 39: egfr gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 174 Grafik 40: egfr gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 174 Grafik 41: olıg2 gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 175 Grafik 42: olıg2 gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 175 Grafik 43: cox-2 gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 176 Grafik 44: cox-2 gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 176 Grafik 45: htert gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 177 Grafik 46: htert gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 177 Grafik 47: dmbt1 gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 183 Grafik 48: dmbt1 gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 183 XII
17 Grafik 49: mgmt gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 184 Grafik 50: mgmt gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 184 Grafik 51: pıke gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 185 Grafik 52: pıke gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 185 Grafik 53: pten gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 186 Grafik 54: pten gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 186 Grafik 55: rb1 gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 187 Grafik 56: rb1 gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 187 Grafik 57: runx3 gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 188 Grafik 58: runx3 gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 188 Grafik 59: p53 gen kopya sayısı rölatif oranlarının tümör grubu ortalaması ve %95 güven aralığı 189 Grafik 60: p53 gen kopya sayısı rölatif oranlarının hücre hatlarındaki ortalaması ve %95 güven aralığı 189 Grafik 61: gstt1 geninin kopya sayısının gen ekspresyonu ile diffüz astrostom grubunda pozitif ilişkisi 190 Grafik 62: gstt1 geninin kopya sayısının gen ekspresyonu ile glioblastoma grubundaki pozitik ilişkisi 191 Grafik 63: gstt1 geninin kopya sayısının gen ekspresyonu ile meningiom grubundaki pozitik ilişkisi 191 Grafik 64: cox-2 geninin kopya sayısının gen ekspresyonu ile meningiom grubundaki pozitik ilişkisi 192 Grafik 65: rb1 geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının tümör gruplarına göre dağılımı 200 Grafik 66: rb1 geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının hücre hatlarına göre dağılımı 200 Grafik 67: rb1 geni promotör bölgesi metilasyonunda tümör grupalrına göre metile/karışım oranları 201 Grafik 68: rb1 geninin promotör bölgesi metilasyonunda; metile/ karışım oranının hücre hatlarına göre dağılımı 201 Grafik 69: pten geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının tümör gruplarına göre dağılımı 202 Grafik 70: pten geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının hücre hatlarına göre dağılımı 202 Grafik 71: pten geninin promotör bölgesi metilasyonunda; metile/ karışım oranının tümör gruplarına göre dağılımı 203 XIII
18 Grafik 72: pten geninin promotör bölgesi metilasyonunda; metile/ karışım oranının hücre hatlarına göre dağılımı 203 Grafik 73: p53 geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının tümör gruplarına göre dağılımı 204 Grafik 74: p53 geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının hücre hatlarına göre dağılımı 204 Grafik 75: p53 geninin promotör bölgesi metilasyonunda; metile/ karışım oranının tümör gruplarına göre dağılımı 205 Grafik 76: p53 geninin promotör bölgesi metilasyonunda; metile/ karışım oranının hücre hatlarına göre dağılımı 205 Grafik 77: runx3 geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının tümör gruplarına göre dağılımı 206 Grafik 78: runx3 geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının hücre hatlarına göre dağılımı 206 Grafik 79: runx3 geninin promotör bölgesi metilasyonunda; metile/ karışım oranının tümör gruplarına göre dağılımı 207 Grafik 80: runx3 geninin promotör bölgesi metilasyonunda; metile/ karışım oranının hücre hatlarına göre dağılımı 207 Grafik 81: mgmt geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının tümör gruplarına göre dağılımı 208 Grafik 82: mgmt geninin promotör bölgesi metilasyonunda; metile/ metile olmayan oranının hücre hatlarına göre dağılımı 208 Grafik 83: mgmt geninin promotör bölgesi metilasyonunda; metile/ karışım oranının tümör gruplarına göre dağılımı 209 Grafik 84: mgmt geninin promotör bölgesi metilasyonunda; metile/ karışım oranının hücre hatlarına göre dağılımı 209 Grafik 85: pık3ca gen ekspresyon pozitifliği sağkalım eğrisi 216 Grafik 86: mdm2 gen ekspresyon pozitifliği sağkalım eğrisi 217 Grafik 87: egfr gen ekspresyon pozitifliği sağkalım eğrisi 217 Grafik 88: gstt1 gen ekspresyon pozitifliği sağkalım eğrisi 218 Grafik 89: olıg2 gen ekspresyon pozitifliği sağkalım eğrisi 18 Grafik 90: cox-2 gen ekspresyon pozitifliği sağkalım eğrisi 219 Grafik 91: htert gen ekspresyon pozitifliği sağkalım eğrisi 219 Grafik 92: dmbt1 tümör süpresör gen kopya sayısı aberasyonunun sağkalım eğrisi 221 Grafik 93: mgmt tümör süpresör gen kopya sayısı aberasyonunun sağkalım eğrisi 211 Grafik 94: pıke tümör süpresör gen kopya sayısı aberasyonunun sağkalım eğrisi 222 Grafik 95: pten tümör süpresör gen kopya sayısı aberasyonunun sağkalım eğrisi 222 Grafik 96: rb1 tümör süpresör gen kopya sayısı aberasyonunun sağkalım eğrisi 223 Grafik 97: runx3 tümör süpresör gen kopya sayısı aberasyonunun sağkalım eğrisi 223 Grafik 98: p53 tümör süpresör gen kopya sayısı aberasyonunun sağkalım eğrisi 224 XIV
19 Grafik 99: onkogen ekspresyon pozitifliği sağkalım eğrisi 225 Grafik 100: tümör süresör gen ekspresyon pozitifliği sağkalım eğrisi 227 XV
20 Kısaltmalar: β-aktin : Beta-aktin µl :Mikrolitre µm :Mikromolar µmol : Mikromol 0 C :Santigrat derece aa : Aminoasit Ca +2 : Kalsiyum +2 iyonu cdna : Komplementer deoksiribonukleik asid cdna : Komplementer DNA CO 2 : Karbondioksit COX 2 : Siklooksigenaz 2 CP : Geçiş noktası Daoy : Desmoplastik serebellar medulloblastoma hücre hattı Dk : Dakika DMBT1 : Malin beyin tümörlerinde silinmiş 1 geni DMSO : Dimetil sülfoksit DNA : Deoksiribonukleik asit DNAse : Deoksiribonukleaz EGFR : Epidermal Büyüme Faktörü Reseptörü F : Forward- İleri Primer FBS : Fötal sığır serum GAPDH : Gliseraldehit-3-fosfat dehidrogenaz gr : Gram GSTT1 : Glutatyon S-transferaz teta 1 htep1 : İnsan telomeraz ilişkili protein 1 htert : İnsan telomeraz revers transkriptazı htr : İnsan telomeraz RNA IC 50 : Letal doz 50 LN-18 : Glioblastoma; glioma hücre hattı LOH : Heterozigozite Kaybı M : Metile M : Molar MDM2 : Transforme olmuş 3T3 hücre double minute 2, p53 bağlayıcı protein Mg+ : Magnezyum MGMT : O 6-metilguanin-DNA metiltransferaz Mix : Karışım ml : Mililitre mm : Milimolar mrna : mesajcı ribonukleik asid MS-PCR : Metilasyon spesifik PCR ng :Nano gram nm : Nanomolar nmol : Nanomol OD : Absorbans değeri Olig2 : Oligodendrosit transkripsiyon faktörü 2 XVI
21 Ort : Ortalama P : Prob PBS : Fosfat tampon tuzu PCR : Polimeraz zincir reaksiyonu PIK3CA : Fosfoinosit 3 kinaz, katalitik, alfa polipeptit PIKE : Fosfotidilinositol 3-kinaz arttırıcı (CENTG1) PTEN : Fosfataz ve tensin homoloğu (MMAC 1) R : Reverse- Geri Primer RB1 : Retinoblastoma RNA : Ribonukleik asid Rpm : Dakikada devir sayısı RT-PCR : Revers transkriptaz polimeraz zincir reaksiyonu RUNX3 : Runt ile ilişkili transkripsiyon faktörü 3 SD : Standart Sapma Sn : Saniye SSS : Santral Sinir Sistemi TAMRA : 6-karboksi-tetrametil-rodamin TdT : Terminal deoksinukleotit transferaz TRAP :telomerik tekrar amplifikasyon protokolü U-118 MG : Glioblastom, astrositom hücre hattı U-87 MG : Glioblastom, astrositom hücre hattı UM : Metile olmayan UV : Ultraviyole WHO: : Dünya Sağlık Örgütü WI-38 : İnsan akciğer fibroblast hücre hattı XVII
22 BÖLÜM I 1.1 GİRİŞ Primer beyin tümörleri, hastalıkların oldukça heterojen bir grubunu oluşturmaktadır. Bu nedenle beyin tümörlerinin heterojen kategorilerini, klinik olarak homojen alt guruplara karakterize edebilmek için uygun sınıflama ölçütlerine ve moleküler tümör işaretlerine gereksinim gittikçe artmaktadır. Bu ve diğer kavramlar beyin tümörlerinin etiyolojisinde hastalığın tek bir nedeni olmadığı fikrini güçlendirmektedir. Tümörlerin moleküler karakterizasyonları, alt tiplerinin benzer moleküler lezyonlara sahip bir gurup hücreden oluşması, tümörlerin açıklanmasında araştırıcılara yardım edebilmektedir. Genlerin ya da proteinlerin dizilerinin incelenmesi, tümör etiyolojisinin lezyonlara ya da tanısal önemine göre daha homojen gruplara ayrılarak sınıflandırılmasına büyük katkı sağlayabilir. Bu moleküler bilgiyi yorumlarken karşılaşılacak en önemli zorluk, hangi değişimlerin etiyoloji açısından öneminin ilk belirtileri ve hangilerinin klinik ve prognostik sonuçları doğuran daha sonraki belirtiler olduğunun tespit edilmesidir. Genetik ve moleküler epidemiyolojik metotlar tümörün moleküler değişimi hakkında karmaşık bilgiler edinmemize yardımcı olmaktadır. Anlamlı tümör işaretleyicilerin ve duyarlılık genlerinin keşfine yönelik çalışmalar, primer beyin tümörleri ve nedenleri hakkında moleküler epidemiyolojik araştırmalar için nörocerrahları, onkologları, patologları, epidemiyologları ve tıbbi biyologları bir araya getirmektedir. Primer beyin tümörleri olası ekzojen ve endojen olaylardan kaynaklanmaktadır. Beyin malignansileri yıkıcı hastalıklardır ama bunların
23 sebeplerini açıklamak için yapılan çalışmalar umut vericidir ve moleküler yaklaşımlar nöro-onkogenez hakkındaki yeni kavramları anlamamızı sağlamaktadır. Gelecekte, beyin tümörlerinin sağkalım oranı, spesifik tanısal genetik ve ekspresyon profillerine bağlı olarak moleküler tabanlı tedavi opsiyonlarının kullanımıyla uzatılması ümit edilmektedir. Çalışmada amaçlanan tümör süpresör ve onkogenlerin gen ve transkript düzeyinde kantite edilmesi ve aralarındaki ilişkinin gösterilmesi literatürde ilk olmakla birlikte çalışmanın özgünlüğünü oluşturmaktadır. Ayrıca çalışma kapsamındaki tümör süpresör genlerin epigenetik düzenlenmelerinin kantitasyonu da ilk olmaktadır. Tanı konduktan sonra sağkalımın ilk yıl için % 20 gibi oldukça düşük bir oranda olmasından yola çıkarak, olgulardaki tümör süpresör ve onkogenlerin profilleri ile hastalıksız sağkalım arasındaki korelasyon belirlenerek optimum tedavi protokollerin saptanması ile tedavi ücretleri azaltılarak ekonomiye ve olguların olası yaşam süre ve kalitelerini arttırarak insani değerlere katkıda bulunulması amaçlanmıştır. Çalışma kapsamında hedeflenen başlıca çıktılar: a) beyin tümörü fenotipine neden olan tümör süpresör ve onkogen profillerinin saptanması, b) ilgili genlerin genetik ve epigenetik değişimlerin kantitasyonuyla kanserin prognozu arasındaki korelasyonun belirlenmesi c) yeni olgularda bu korelasyon ile prognozunu belirlemede prediktif olarak kullanarak daha ekonomik ve optimum tedavinin gerçekleşmesi d) kesin tanı almamış olgularda tanıya moleküler bir yaklaşım getirilmesi e) transkripsiyon ile translasyon arasındaki korelasyon ile tümör 2
24 tedavisinde yeni moleküler yaklaşımlar sağlanması f) kanser fenotipi ile ilgili genlerin genetik ve epigenetik değişimlerinin tanı ve prognozda katkı sağlamasıdır 1.2. GENEL BİLGİLER Beyin (Encephalon) Beyin, cavitis cranii içerisinde yer alır ve foramen magnum da medulla spinalis ile devam eder. Üç meninges ile çevrilidir: dura mater, arachoidea mater ve pia mater. Bu örtüler omuriliğin ilgili meningesleriyle devam eder. Beyin üç temel bölüme ayrılır. Bunlar omurilikten yukarıya doğru rhombencephalon, mesencephalon ve prosencephalon. Rhombencephalon medulla oblongata, pons ve serebellum olmak üzere üç alt bölüme ayrılabilir. Prosencephalon da merkez bölümünü oluşturan diencephalon (ara beyin) ve cerebrum a ayrılır. Beyin sapı (medulla oblongata, pons ve mesencephalon için ortak terim=truncus cerebri) serebral hemisferler ve serebellum uzaklaştırılıdığında beynin geriye kalan bölümüdür. Medulla spinalis ten farklı olarak beyin iç kısmında beyaz cevher, dışta bunu saran gri cevherden oluşur. Ancak bazı önemli gri cevher kitleleri beyaz cevherin derininde bulunur. Örneğin serebellum içerisinde gri serebellar çekirdekler (nuclei cerebellares) ve serebrum içerisinde gri nuclei thalami, nucleus caudatus ve nucleus lentiformis yerleşmiştir (317). 3


25 Lobus frontalis Polus frontalis Fissura longitudinalis cerebri Sulcus precentralis Gyrus precentralis Hemispherium cerebri Sulcus centralis Gyrus postcentralis Lobus parietalis Sulcus postcentralis Lobus occipitalis Lobus frontalis Hemispherium cerebri Lobus temporalis Mesencephalon Pons Serebellum Polus occipitalis Fissura longitudinalis cerebri Tractus olfactorius Chiasma opticum N. oculomotorius Pyramis Medulla oblongata Oliva Şekil 1: Beynin üstten ve alttan görünüşü (Snell den değiştirilerek alınmıştır, 317) 4
26 Beyin Tümörleri Tümör etiyolojisi sinir sisteminin her kesiminde rastlanabilecek bir olaydır. Beyin, Medulla spinalis ve periferik sinirleri etkileyecek şekilde klinik tablolara yol açabilir. Her sistem veya topografik vücut bölgesinde olduğu gibi sinir sisteminin tümörleri de bir sınıflandırma doğrultusunda incelenebilir. Böyle bir sınıflandırma çabası ilk kez 19. Yüzyılın sonlarında Wirchow tarafından gösterilmiş ve bugün de kullandığımız (glioma-nörinoma) gibi isimler türetilmiştir. Hücre niteliğini esas alan bu ilk çalışma sonraki yıllarda geliştirilmiş, Hortega, Bailey-Cushing, Kernohan, Zülch isimli çalışmacılar kendi adlarıyla anılan sınıflamalar bildirmişlerdir. Embriyogenez ve sitolojiye dayanan değerlendirmeler üzerine halen tartışmalar yapılmakta ve ayrı tanımlamaların eşdeğerliği aranmaktadır. Klinikçiler, bir sınıflandırmanın biraz da anatomik lokalizasyonu, klinik antiteleri ve prognozu yansıtmasını ister. Nitekim klinik yazarlar bu doğrultuda derlemeler yapmaktadır. Bu çabalara kalırsa, tüberküloma ve parazit kisti gibi aslında enfeksiyon olan bazı etiyolojiler de tümör terimi ile anılmaktadır. Yada, beyin tümörü başlığına kranium ve hipofiz gibi aslında sinir sistemi ile ilgisiz dokuların tümörleri de sokulmaktadır. Pratik yaklaşım, anatomik lokalizasyon ve semptomatolojiyi esas almak ve ayrıntıları patolojik özelliklere göre incelemektedir (165) Epidemiyoloji Beyin kanseri tüm kanser tiplerinin yaklaşık olarak %1,4 ünü ve ölümle sonuçlanan kanserlerin de %2,3 ünü oluşturmaktadır. Primer serebral malignansilerin insidansı tüm toplum içerisinde 4 ve 10/100,000 arasında 5
27 değişmektedir. Bu insidans yaşla (12 yaşına kadar 4/100,000; 35 yaşına kadar 6/100,000; 55 yaşına kadar 18/100,000; 75 yaşına kadar 70/100,000) yılında, 35,000 in üzerinde Amerikalı ya (yaklaşık 6/100,000) beyin tümörü tanısı konmuştur. Sınıflandırılan şartlarda bulunan gruplardan elde edilen verilere göre yıllık ölüm oranı 13,000/yıl dır. Günümüzde, gelişmiş tanı yöntemleriyle, yaklaşık 16,800 beyin tümörlü vakaya her yıl malin tanısı konmuştur (10). Ancak, benin tanısı konan ve tedavi edilebilir durumda olan vakalar, normal bir yaşam için önemli olan normal beyin fonksiyonları ile çatışmaktadır (10). Olgular için süre gelen amansız manzara, tedavi edilebilen düşük dereceli pediatrik kanserlerin bile tahrip edici etkisi ve diğer benin rahatsızlıklar beyin kanseri araştırmalarına yeni bir hız vermiştir. İki tip olan; tanımlayıcı ve analitik epidemiyolojik çalışmalar, beyin tümörlerine ilişkin yapılan araştırmalarda son zamanlarda artan bir hal almıştır. Tanımlayıcı çalışmalar yaş, cinsiyet ve coğrafik bölge gibi hasta demografisini ve histolojik tümör tipini kategorize ederek beyin tümörlerine ilişkin insidansı, mortaliteyi ve yaşama oranını karakterize etmektedir. Analitik epidemiyolojik çalışmalar toplulukta bulunan, belirli karakteristikleri ve geçmişleri olan ve olmayan bireylerde beyin tümörü riskini karşılaştırır, kanser gelişimi sırasında belirebilecek risk faktörlerini araştırır. Beslenme, sigara ve alkol kullanımı, mesleki problemler, radyasyon, alerji, kafa travmaları ve aile öyküsü gibi birçok risk faktörünün beyin tümörü oluşumundaki rolleri araştırılmaktadır. Son yıllarda, genlerin çevresel etkileşimlerinde olduğu gibi, karsinojen metabolizması ve DNA tamirine ilişkin genlerdeki kalıtılan polimorfizmler 6
28 büyük ilgi çekmektedir. Beyin tümörlerinin göreceli seyrekliği büyük toplumlar üzerinde yapılması planlanan çalışmaları zorlaştırmaktadır, bu nedenle, bu analitik çalışmaların çoğu genellikle olgu-kontrol yaklaşımını kullanmaktadırlar. Artan bir şekilde gelişen, epidemiyolojik beyin kanseri çalışmalarına rağmen, bireysel risk faktörlerinin doğası ve ağırlığı üzerine oldukça az görüş birliği bulunmaktadır. Çalışma dizaynındaki, popülasyondaki veri kaynakları ve sınıflandırmadaki varyasyonlar araştırmalar üzerine gölge düşürmektedir. Çalışmalar, çalışma konularının seçimi, konuların iyi bir örnek olup olmadığına karar verilmesi ve kontrol gruplarının seçimi ve tanımlanması için olanlar gibi metodoloji açısından farklıdır. Çalışmalar, proksi ve geçmiş bilgileri üzerine güvenilirlikleri açısından değişiklik gösterirler ve bunların doğruluk ve bütünlük standartları örtüşmelidir. Ayrıca, bu çalışmalarda, primer beyin tümörlerinin heterojenitesinden kaynaklanan hisyolojik tanıdaki tutarsızlıklar, tanımlamalar ve gruplaşmalar gibi temel klasifikasyon problemleri sorun olmamalıdır. Son olarak, 2007 yılında, Dünya Sağlık Örgütü (WHO) tümör klasifikasyonu güncelleştirilmiştir Olguların Yaş ve Cinsiyeti Farklı etiyolojik faktörleri olan beyin tümörlerinin farklı histolojik tiplerinin olma olasılığı yaş dağılımı ve tümörün bölgesi ve histolojisindeki farklılıklardan kaynaklandığı ileri sürülmektedir. Tüm primer beyin tümörleri için, olgunun ortalama yaşı 54 civarında olmasına rağmen, her histolojik kategori için önemli bir vasyasyon bulunmaktadır. Örneğin, glioblastom ve meningiomlar için ortalama yaş 62 dir (212). Meningiomlarda, insidans, 85 ve üzeri yaşta olan olgular için azalmanın 7
29 dışında, yaş ile artmaktadır. Bunun tersine, astrositom ve glioblastomlarda insidans yaşları arasında zirveye ulaşırken oligodendrogliomlarda insidans yaşları arasında zirveye ulaşmaktadır. Bu varyasyonları bazıları farklılaşan tanısal uygulamaları ve farklı yaş gruplarına yapılan girişimi yansıtabilir. Yaşla ilişkili tümör insidansının çoğu malin transformasyon için gerekli olan etkiye maruza kaldıkları süreden, klinik hastalık ya da gelişen yaşla birlikte zayıf immun gözetim oluşturmak için gerekli olan genetik değişikliklerin sayısından sorumlu tutulmaktadır. Beyin tümörlerinin şaşırtıcı ve tam olarak açıklanamayan özellikleri küçük çocuklarda görülen insidansta pik yapmaktadır ve bunlar pediatrik tümör olan primitif nöroektodermal orijinli tümörlere tamamen yorulamamaktadır. Genellikle, erkeklerde bayanlara göre daha yüksek oranda primer beyin tümörü gözlenir. Ancak, bayanları erkeklere göre %80 oranında daha fazla etkileyen meningiomalar ve erkek ve bayanları eşit oranda etkileyen kraniyal ve spinal sinir tümörleri bu durumun dışında kalmaktadır. Gliomlar erkekleri bayanlara göre %40 daha fazla etkilerler (325). New York da yapılan bir çalışmaya göre glioblastomlarda cinsiyet farklılıkları adet görme yaşıyla birlikte görülmeye başlıyor, menapoz döneminde pik yapıyor ve daha sonra gittikçe azalıyor. Bu durumun, hormonların koruyucu etkisinden kaynaklandığı düşünülmektedir (213). Beyin tümörlerinin yayılımı ve sebeplerine ilişkin kapsamlı bir çalışma düzenli bir şekilde gözetim altında tutulan yaş ve cinsiyet farklılıkları için bir açıklama içermelidir. İnsidans daha iyi ve daha tutarlı bir tanının sonucu olarak daha doğru bir şekilde açıklandıkça, tümörlerin moleküler klasifikasyonundaki dramatik ilerleyişi tümörlerin homojen alt gruplarının etiyolojik tanımlanması olasılığını arttırmaktadır. Güncel olan genlerin ivmeli 8
30 karakterizasyonu, hangilerinin beyin tümörlerine ya da etiyolojik çevresel ajanlara olan duyarlılığı ya da direnci arttırabileceğine karar verme fırsatını da yaratmaktadır. Karsinogenezise olan spesifik ilginin nedeni, hücre bölünmesini aktive ederek karsinogenezisi başlatan proto-onkogenler ve tümör gelişimini ve progresyonunu inhibe eden süpresör genlerdir (353). Bu gibi genler hastalığın progresyonunda ve radyasyona ya da ilaç müdahalesine olan duyarlılıkta (ya da dirençte) rol oynadıkları için çalışmalardan elde edilecek bilgi uygulanabilir bir önleme stratejisi oluşumunda kullanılabilir Beyin Tümör Tiplerinin Histolojisi ve Moleküler Biyolojisi Primer beyin tümörleri histolojik ve yerleşim bölgesi olarak sınıflandırılmıştır. Ancak, bu sınıflandırma, sinir sistemi hücrelerinin, birçok beyin tümörü tipinde sıklıkla gözlemlenen hücre tiplerinin karışımı ile sonuçlanan kanserli kanserleşebilme yeteneğinden ötürü oldukça komplikedir. Beyin tümör tipleri, invasivliklerine ve malinitelerine göre WHO tarafından kategorize edilmişlerdir. Daha sık gen mutasyonu gösteren daha invaziv formlar daha yüksek değerler verirler. Beyin tümörleri üzerine yapılan son çalışmalardan elde edilen veriler aşağıda özetlenmiştir Sinir Sistemi Tümörlerinin Sınıflandırılması Nöroepiteliyal dokunun tümörleri Astrositik tümörler Oligodendroglial tümörler Miks gliomalar Ependimal tümörler Koroid pleksus tümörleri Belirsiz orijinli Glial tümörler Nöronal ve miks nöronal-glial tümörler 9
31 Nöroblastik tümörler Pineal parenkimal tümörler Embriyonel tümörler Periferal sinirlerin tümörleri Schwannoma Nörofibroma Perinörioma Malin periferal sinir kılıfı tümörleri Meningelerin tümörleri Meningoteliyal hücrelerin tümörleri Mesenşimal, non-meningoteliyal tümörler Primer melanositik lezyonlar Belirsiz histogenez tümörleri Lenfomalar ve hemopoietik neoplazmlar Germ hücre tümörleri Sellar bölge tümörleri Metastatik tümörler Dünya Sağlık Örgütü (WHO) Derecelendirmesi Histolojik derecelendirme ile amaçlanan tümörün biyolojik davranışının önceden kestirilmesini sağlamaktır. Klinik uygulamalarda tümör derecesi seçilecek tedavi modalitesinin belirlenmesinde, özellikle adjuvan radyoterapi ve özel kemoterapötiklerin seçilmesinde önem arz etmektedir. WHO nün tümör sınflandırmasında derecelendirme, çok çeşitli histolojik özelliğe sahip tümörlerin malignansi ölçütüdür. Derece 1: Düşük proliferasyon (çoğalma) potansiyeline sahip ve cerrahi olarak çıkarılmasını takiben kür şansı bulunan tümörlerdir. Derece 2: Genel olarak infiltratif (sızma) tabiyata sahip ve düşük prolifratif potansiyele sahip olmasına karşın sıklıkla tekrarlayan tümörlerdir. Bazı tip 2 tümörler de yüksek dereceli tümörlere dönüşme eğilimli mevcuttur örneğin düşük dereceli astrositom, anaplastik astrositoma ve glioblastoma dönüşebilmektedir. 10
32 Derece 3: Genellikle histolojik olarak malignansi bulguları gösteren lezyonlar için kullanılır (nükleer atipi, aktif bir mitotik aktivite). Çoğunlukla derece 3 tümörlü hastalar adjuvan radyoterapi ve\veya kemoterapi görmektedirler. Derece 4: Sitolojik olarak malin, mitotik olarak aktif, nekroz eğilimi olan neoplazilerdir. Tipik olarak hızlı bir preoperatif ve postoperatif hastalık gelişimi söz konusudur ve ölümcül bir seyir gösterirler. NÖROEPİTELYAL DOKU TÜMÖRLERİ ICD-0/Davranış Astrositik Tümörler WHO derece /1(*) Pilositik Astrositom derece I 9425/3(**) Pilomiksoid Astrositom derece II /1 Subependimal Dev Hücreli Astrositom derece I /3 Pleomorfik Ksantoastrositom derece II /3 Diffüz Astrositom derece II 9420/3 Fibriller Astrositom 9411/3 Gemistositik Astrositom 9410/3 Protoplazmik Astrositom /3 Anaplastik Astrositom derece III /3 Glioblastom derece IV 9441/3 Dev Hücrell Glioblastom derece IV 9442/3 Gliosarkom derece IV /3 Gliomatozis Serebri Oligodendroglial Tümörler /3 Oligodendrogliom derece II /3 Anaplastik Oligodendrogliom derece III Oligoastrositik Tümörler /3 Oligoastrositom derece II /3 Anaplastik Oligoastrositom derece III Ependimal Tümörler /1 Subependimom derece I /1 Miksopapiller Ependimom derece I /3 Ependimom derece II 9391/3 Sellüler 9393/3 Papiller 9391/3 Berrak Hücreli 9391/3 Tanisitik /3 Anaplastik Ependimom derece III 11
33 Koroid Pleksus Tümörleri /0 Koroid Pleksus Papillomu derece I /1 Atipik Koroid Pleksus Papillomu derece II /3 Koroid Pleksus Karsinomu derece III Diğer Nöroepitelyal Tümörler /3 Astroblastom derece I /1 Üçüncü Ventrikülün Kordoid Gliomu derece II /1 Angiosentrik Gliom derece I Nöronal ve Mikst Nöronal-Glial Tümörler /0 Serebellumun Displastik Gangliositomu (Lhermitte-Duclos) /1 Desmoplastik İnfantil Astrositom / Gangliogliom derece I /0 Disembryoplastik Neuroepitelyal Tümör derece I /0 Gangliositom derece I /1 Gangliogliom derece I /3 Anaplastik Gangliogliom derece III /1 Santral Nörositom derece II /1 Ekstraventriküler Nörositom derece II /1 Serebellar Liponörositom derece II /1 Papiller Glionöronal Tümör derece I /1 V. Ventrikülün Rozet Oluşturmuş Glionöronal Tümörü derece I /1 Paragangliom derece I Pineal Bölge Tümörleri /1 Pineositom derece I /3 İntermediyer Diferansiyasyon Gösteren Pineal Parankimal Tümör derece II, III /3 Pineoblastom derece IV /3 Pineal Bölgenin Papiller Tümörü derece II, III Embriyonal Tümörler /3 Medulloblastom derece IV 9471/3 Desmoplastik/Nodüler Medulloblastoma 9471/3 Şiddetli Nodülarite Yapan Medulloblastom derece IV 9474/3 Anaplastik Medulloblastom derece IV 9474/3 Büyük Hücreli Medulloblastom /3 SSS Primitif Nöroektodermal Tümör derece IV 9500/3 SSS Nöroblastom 9490/3 SSS Ganglionöroblastom 9501/3 Medulloepitelyom 9392/3 Ependimoblastom /3 Atipik Teratoid/Rabdoid Tümör derece IV 12
34 Kranyal ve Paraspinal Sinirlerin Tümörleri /0 Schwannom (Nörilemmom, Nörinom) derece I 9560/0 Sellüler 9560/0 Pleksiform 9560/0 Melanotik /0 Nörofibrom derece I 9550/0 Pleksiform 43 Perinörioma derece I, II, III 9571/0 Perinöriom, NOS 9571/3 Malin Perinöriom 44 Malin Periferal Sinir Kılıfı Tümörleri (MPNST) derece II, III, IV 9540/3 Epitelioid MPNST 9540/3 Mezenkimal Diferansiyasyon Gösteren MPNST 9540/3 Melanotik MPNST 9540/3 Glandüler Diferansiyasyon Gösteren MPNST Meninkslerin Tümörleri Meningotelyal Hücrelerin Tümörleri /0 Meningioma derece I 9531/0 Meningotelyal 9532/0 Fibröz (Fibroblastik) 9537/0 Transisyonel (Mikst) 9533/0 Psammomatöz 9534/0 Angiomatöz 9530/0 Mikrokistik 9530/0 Sekretuar 9530/0 Lemfoplasmositten zengin 9530/0 Metaplastik 9538/1 Kordoid 9538/1 Berrak Hücreli 9539/1 Atipik derece II 9538/3 Papiller 9538/3 Rabdoid 9530/3 Anaplastik (Malin) derece III Mezenkimal Tümörler /0 Lipom /0 Angiolipom /0 Hibernom /3 Liposarcom /0 Soliter Fibrös Tümör /3 Fibrosarkom /3 Malin Fibröz Histiositom /0 Leiomyom /3 Leiomyosarkom /0 Rabdomyom 13
35 /3 Rhabdomyosarkom /0 Kondrom /3 Kondrosarkom /0 Osteom /3 Osteosarkom /0 Osteokondrom /0 Hemangiom /1 Epithelioid Hemangioendotelyom /1 Hemangioperisyitom derece II /3 Anaplastik Hemangioperisitom derece III /3 Angiosarkom /3 Kaposi Sarkomu /3 Ewing Sarkom - PNET Primer Melanositik Lezyonlar /0 Diffüz Melanositozis /1 Melanositom /3 Malin Melanom /3 Meningeal Melanomatozis Meninkslerle İlişkili Diğer Tümörler /1 Haemangioblastom derece I Lenfomalar ve Hemapoietik Tümörler /3 Malin Lenfomalar /3 Plazmositom /3 Granülositik Sarkom Germ Hücre Tümörleri /3 Germinom /3 Embriyonal Karsinom /3 Yolk Sak Tümörü /3 Koriokarsinom /1 Teratom 9080/0 Matür 9080/3 İmmatür 9084/3 Malin Transformasyon GösterenTeratom /3 Mikst Germ Hücre Tümörleri Sellar Bölge Tümörleri /1 Kraniofaringiom derece I 9351/1 Adamantinomatöz 9352/1 Papiller /0 Granüler Hücreli Tümör derece I /1 Pituisitom derece I /0 Adenohipofizin İğsi Hücreli Onkositomu derece I 14
36 Metastatik Tümörler (*) International Classification of Diseases for Oncology (ICD-O) {614A} ve the Systematized Nomenclature of Medicine ( Morfoloji kodları. Davranış: /0 benign tümörleri, /3 malin tümörleri ve /1 borderline veya davranışı bilinmeyen tümörleri göstermektedir (338) Gliomlar Gliomlar sinir sisteminin glial bileşenlerinden oluşurlar ve hücreleri nöronlar ve beyin sıvısı arasında bir ara yüzey oluşturur. En sık rastlanan primer beyin tümörleridir ve 60 lı yaşlarda insidanstaki artışla birlikte sentral sinir sistem neoplazmlarının %40 ından fazlasına neden olmaktadır (152). Gliomlar astrosit, oligodendrosit ya da ependimal hücrelerinden köken almalarına rağmen aralarında başlangıç dönemlerinde içerdikleri genleri yansıtan önemli değişiklikler vardır (153). Genellikle, glial tümörler orijinlerindeki varsayılan hücre tiplerine göre adlandırılırlar: Bunlar; astrositomlar, oligodendrogliomlar ve ependimomlardır Astrositomlar Astrositomlar beyin tümörlerinin çoğunluğunu oluşturmaktadırlar. Sentral sinir sistemindeki nöronlara yaygın yapısal ve fizyolojik destek sağlayan her yerde yaygın olarak bulunan yıldız-şekilli hücrelerdir. Astrositik tümörler SSS de geniş oranda bulunan lokalizasyon, yaş ve cinsiyet dağılım, büyüme potansiyeli, istilacılık genişliği, morfolojik özellikleri, progresyon eğilimleri ve klinik seyirleri farklılıklar sergileyen neoplazmlardır. Bu farklılıkların, transformasyon işlemi sırasında kazanılmış genetik değişimlerin sekansı ve tipini yansıttığının kanıtları artmaktadır. 15
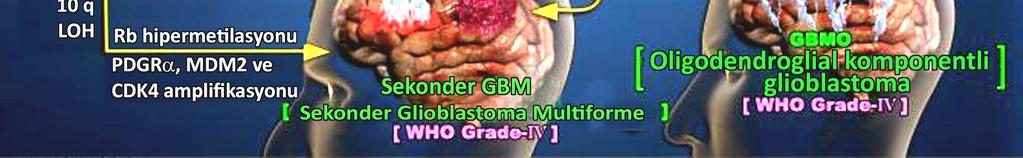
37 Dünya Sağlık Örgütü, astrositomların klinikopatolojik alt tiplerini aşağıdaki gibi ayırt etmektedir. a. WHO derece I, veya pilositik astrositom, çocuklarda rastlanan en yaygın beyin tümörüdür ve esasen pediatrik bir tümördür ve nadiren neoplastik transformasyon geçirmektedir. Lokalizasyona bağlı olarak en benign astrositomlar, vital duyusal fonksiyonları engelleyebilmekte ve sıklıkla tam bir rezeksiyon ardından rekürrens sergilemektedirler. b. WHO derece II, veya fibriler astrositomlar, tüm gliomların %25 ini oluşturmakta ve doğal olarak nüfuz ederler (infiltratiftirler). Agresif histolojik özellikleri olmamasına rağmen, erişkinlerdeki düşük-dereceli astrositomlar, hastaların büyük çoğunluğunda öldürücüdürler. c. WHO derece III, veya anaplastik malin astrositomlar, yüksek oranda malin gliomlardır ve artmış glioblastoma gelişimi eğilimine sahiptirler. d. WHO derece IV, difüz astrositom veya glioblastoma multiforme, son derece malin beyin tümörleridir ve tipik olarak erişkinleri etkilerler. Bu gliom tipleri kötü prognoza sahiptirler, kısmen, kötü olarak tanımlanan tümör, hızla beynin diğer bölgelerine yayılırlar. Bunlar en yaygın intrakranial neoplazmlardır ve primer beyin tümörlerinin %60 ını açıklarlar. Astrositomlardaki genetik değişimler, artan malignansi ve klinik derecenin nonrandom (rastgele olmayan) bir dizi genetik değişikliğin farkına varılmasını sağlamıştır (166). Yaygın kromozomal değişimlerin, çeşitli genlerin ekspresyonları ve/veya fonksiyonlarındaki değişiklikler arasındaki ilişkilerinden kaynaklandığı 16
38 gözlenmiştir. Örneğin, kromozom 17p de bulunan p53 genindeki mutasyonlar, astrositik tümörlerin %40 ının tüm derecelerinde rapor edilmiştir. Bu mutasyonlar primer olarak, çocuklara göre genç erişkinlerde daha çok ortaya çıkmaktadır (109). Düşük dereceli gliomlarda yüksek derecelilere göre daha sık rastlanan bir gen kaybı veya değişikliği ise siklin-bağımlı kinaz N2 (CDKN2) veya p16 geninde gözlenir. Kromozom 9p de lokalize CDKN2 geninin her iki kopyasının da delesyonu yüksek dereceli astrositomlarda yaygındır (141). Delesyonlara ek olarak, p16 genindeki yeniden düzenlenmeler de rapor edilmiştir. Kromozom 10 un (çeşitli tümör süpresörleri barındıran) delesyonları (131), yüksek dereceli astrositik tümörlerde sıklıkla ortaya çıkmaktadır. 10q23 teki heterozigisite kaybı glioblastomların yaklaşık %70 inde rapor edilmiştir. 10q kromozomunda lokalize bir tümör süpresör olan MMAC1 veya fosfataz tensin homoloğu (PTEN), glioblastomların %40 ında mutasyona uğramakta, fakat düşük dereceli gliomlarda nadiren mutasyon görülmektedir. Sonuç olarak, MMAC1 düşük dereceli ve yüksek dereceli glioma progresyonunda önemli rol oynar (28). 17








39 Şekil 2: Glioma patogenezi (123) 18
40 Gliomlardaki bazı kromozomal değişiklikler, sıklıkla artmış ve değişime uğramış hücre proliferasyonu ile sonuçlanan tümör süpresör genlerin kaybı ve onkogenlerin aktivasyonlarını içermektedir. Epidermal büyüme faktör reseptör (EGFR) geni, malin astrositomlarda en sık amplifiye olan gendir. EGFR proteini, epidermal büyüme faktörünün reseptörü ve astrositler için önemli bir proliferatif uyarıcıdır (171). Mutasyona uğramış bir EGFR alelinin amplifikasyonu glioblastomların yaklaşık olarak üçte birinde bulunmakta fakat, düşük dereceli astrositomlarda gözlenmemektedir (125) Oligodendrogliomlar Oligodendrogliom, sinir hücrelerinin aksonlarını çevreleyen yağı üreten hücreler olan oligodendrositlerden gelişmektedir. Bu tip bir tümör normal koşullarda serebrumda ortaya çıkmakta ve çocuklara nazaran erişkinlerde, bayanlara göre erkeklerde daha yaygın olarak gözlenmektedir. Oligodendrogliomlar, tüm glial tümörlerin %5-12 sini ve tüm intrakranial tümörlerin %5-7 sini meydana getirmektedirler. Yavaş büyüme eğilimleri ve X-ray incelemeleri, bilgisayarlı tomografi ve aynı zamanda histolojik incelemelerde karakteristik kalsifikasyon sergilerler. Oligodendrogliomlar klinik olarak astrositomlara göre daha az agresif olmalarına rağmen; invazivdirler ve serebral spinal sıvı içinden geçebilirler. Metastaz yapma kapasiteleri vardır ve sıklıkla cerrahi olarak alınmaları daha zordur. Buna rağmen, diğer gliomlara göre daha iyi prognoz ve sağkalım sergilerler. Diğer gliomlar gibi, oligodendrogliomlar malignansi ve büyüme hızlarına göre 1 den 4 e kadar derecelendirilirler. 19
41 Oligodendrogliomlar karakteristik olarak 1p ve 19q13 kromozomal bölgelerde kayıp ve daha az sıklıkla ise 9q ve 22. kromozomlarda kayıplar segilerler (219, 278). Hoang-Xuan ve arkadaşları (117), 26 oligodendrogliomun (10 olgu derece II ve 16 olgu derece III) moleküler profilini incelemişler ve en sık rastlanan değişim olarak, 1p ve 19q kromozomal bölgelerinin heterozigosite kaybı olduğunu gözlemlemişlerdir. Bu iki değişim, çok yakından ilişkilidir ve tümörogenezde bu iki lokusun aynı yolakta olduğu izlenimini bırakmaktadırlar. Bu çalışma ayrıca, P16/CDKN2A tümör süpresör geninin homozigot delesyonunun kombinasyonu ile kromozom 10 üzerinde yer alan heterozigosite kaybı (LOH), ve EGFR onkogeninin amplifikasyonunun daha önce rapor edildiğinden daha yüksek oranda var olduğunu göstermiştir (118). İstatistiksel olarak anlamlı bir dışlama ise, bu üç genetik değişim ve 1p/19q daki LOH arasında bildirilmiştir. Bu durum ise oligodendrogliomda en az iki bağımsız genetik altküme olduğunu işaret etmektedir. EGFR amplifikasyonu ve 10q daki LOH daha kısa progresyondan bağımsız sağkalımın (PFS) önemli prediktörleridir, bu nedenle tümörün daha agresif bir formunu karakterize etmekte iken 1p deki LOH daha uzun PFS ile ilişkilendirilmektedir Ependimomlar Bu tümörler, beynin ventriküllerini ve spinal kordun sentral kanalını dolduran ependimal hücrelerden gelişmektedirler. Ependimomlar, serebrosipinal sıvı (CSF) aracılığı ile beyinden spinal korda yayılabilirler ve ventrikülün veya hidrosefalinin göze çarpan şekilde şişmesine neden olurlar. Tüm beyin tümörlerinin %4-6 sını oluştururlar ve çoğunlukla 20 yaşa kadar ortaya çıkarlar. Çocuklarda, ependimomların %30 u 3 yaşından önce ortaya çıkmakta ve erişkinlere göre daha 20
42 agresif seyretmektedirler. Pediatrik ependimomların neredeyse %90 ı intrakranialdir: supratentorial veya posterior fossada lokalizasyon gösterirler ve yalnızca %10 u intraspinaldir. Daha önce söz edilen tümörlerin aksine, düşük dereceli ependimomlar (derece I/II), neroeksen boyunca metastaz geliştirirler. Ependimomlarda en çok tanımlanan genetik değişiklik 17p delesyonları ve monozomi 22 dir (166) Meningiomlar Meningiomlar, beyni çevreleyen meningelerin (beyni ve omuriliği çevreleyen zarlar) endoteliyal kaplama (cover) hücrelerinden köken almakta ve benin tümörler olarak kabul edilmektedirler. Tüm beyin tümörlerinin %10-19 unu oluştururlar. Meningiomlar, kraniyal bazdaki tümörlerin geniş bir oranını oluştururlar. Bundan dolayı; anatomik malignansi terimi bu bölgedeki meningiomları belirtmek için kullanılır (biyolojik malignansinin tersidir). Çocukların %2 sinden azında görülmesine rağmen, meningiomların yaş dağılımı homojendir. Amerika da her yıl en az 3000 yeni meningiom vakası teşhis edilmekte ve 2:1 oranında çoğunlukla bayanlarda erkeklerden daha fazla görülmektedir. Tümörler tüm yaş gruplarında ortaya çıkmakta ama genellikle orta yaşlarda pik yapmaktadır. Yüksek dereceli veya malin meningiomlar, kromozom 1 üzerindeki lokuslardaki delesyonlar ile ilişkilidir ve, daha düşük boyutta ise 6p, 9q, ve 17p üzerindeki delesyonlar ile karakterizedirler (166). Malin meningiomlarda p53 genindeki mutasyonlar da rapor edilmiştir. 21
43 Medulloblastomlar Medulloblastomlar, primitif veya yetersiz gelişmiş hücrelerden köken alan malin tümörlerdir, beyin tümörlerinin %3-5 ini ve çocukluk çağı tümörlerinin %25 ini oluştururlar. Hastalık yaygın olarak 3 ile 8 yaşları arasında ortaya çıkmakta, zaman zaman ise yetişkinlerde de gözlenmektedirler. Bu lezyonların medyan lokalizasyonları ve dördüncü ventrikül ile ilişkileri nedeniyle, bunlara sıklıkla, ventriküler sistem ve nöroeksen metastazları eşlik eder (%25 45). Olguların %5 inde tanı esnasında dahi hâlihazırda metastazlar mevcuttur. Medulloblastomlar en sık rastlanan pediatrik malignansiler olmalarına rağmen, çocukluk çağı medulloblastomlarının uzun-evreli sağkalım sergilemelerinin nedeni hakkında çok az şey bilinmektedir. Medulloblastomun radyasyon ile tedavisi, sekonder malignansilerin gelişimini kapsamaktadır. Medulloblastomlarda kromozom 17p delesyonları yaygındır. Diğer az rastlanan delesyonlar ise 2p, 6q, 10q, 11p, 11q ve 16q da lokalizedir (166) Ganglioglioma Gangliogliomlar, nöronları ve glial hücreleri içeren tümörlerdir ve genellikle temporal loblarda ve serebral hemisferlerde ortaya çıkmaktadırlar. Yüksek oranda yalnızca cerrahi ile veya cerrahi ile kombine radyasyon ile tedavi edilebilinmektedirler. Merkezi gangliomlar çocukların %0,4-8 i ve yetişkinlerin %1 inde ortaya çıkmakta ve cinsiyet veya ırk ile insidansta farklılık sergilemez. Gangliogliomların çoğunluğu, 30 yaşından daha genç hastalarda gözlenir. Olguların % ünde gangliogliomlar, bilgisayarlı tomografide (CT) veya magnetik 22
44 rezonans görüntüleme (MRI) değişimler görülmeden çok uzun bir süre önce oluşan, epileptik nöbet ile açığa çıkmaktadırlar (249). Gangliogliomda görülen genetik değişimler arasında kromozom 9 un kısa kolundaki genetik materyalin kaybı ve kromozom 7 nin bir kısmının veya tümünün aşırı ifadesidir. Tuberoz skleroz 2 (TSC2) genindeki polimorfizmin, sporadik gangliogliomların gelişimine yatkınlık oluşturabileceği yakın zamanda rapor edilmiştir Schwannomlar (Nörilemomlar) Schwannomlar, Schwann hücrelerinden köken alan genellikle benign tümörlerdir ve sıklıkla serebellum yakınında ve duyma ve dengeden sorumlu kraniyal sinirlerde oluşurlar (109). Bu benign tümörler, bayanlarda erkeklere oranla 2 kat daha yaygındır ve çoğu kez 30 ile 60 yaşları arasında teşhis edilirler. İntrakraniyal schwannomlar, primer beyin tümörlerinin yaklaşık %8 ini oluştururlar. En yaygın görülen schwannom, sekizinci kraniyal sinir tümörü akustik nöromdur. Fakat bu tümörler diğer kraniyal sinirlerde ortaya çıkabilirler. Malin schwannom, periferal sinirlerden orijin almakta ve rekürren hastalık ve erken metastazlarla birlikte malin bir seyir izlerler. 1p kayıpları ve 11q kazançları bazı schwannomlarda tespit edilmiş, fakat 22q kaybı haricinde tek başına hiçbir sürekli genetik değişim bu güne kadar schwannomlar ile ilişkili bulunmamıştır. Bu tip değişimler daha önce sistematik olarak araştırılmamıştır Kordomalar Kordomalar embriyonik notokordal kalıntılardan baş gösteren nispeten nadir neoplazmlardır ve intrakraniyal neoplazımların %1 inden azını kapsarlar. Nöroeksen 23
45 boyunca, özellikle gelişimsel olarak daha aktif kraniyal ve kaudal uçlarda, dikkati çekecek bir şekilde sfeno-oksipital, sakrokoksigeal, ve vertebral konumlarda yer alırlar. Kordomaların % ı kafatası kaidesi bölgesinde ortaya çıkarlar. Bu tümörler hâkim olarak yaş aralığında ortaya çıkarlar ve erkeklerde az bir hâkimiyet gösterirler. Tedaviye uyumludurlar fakat 10 ile 20 yılı geçkin bir süreçte inatla nüks ederler. Bu hastalık ile ilgili genetik araştırma yoluna gidilmiştir. Bir çalışmada kordomalarda kromozomal dengesizlikler saptanmış ve tümör süpresör genler veya uyumsuzluk tamiri genleri (1p31 ve 3p14 te lokalize) ve onkogenler (7p36 da lokalize) ile ilgili kordomagenez ile ilgili olabileceğini öne süren veriler elde edilmiştir (294) Nörofibromatozis Nörofibromatozis (NF-1), veya von Recklinghausen hastalığı, 3000 canlı doğumdan 1 inde ortaya çıkmakta (60) ve kalıtımsal model olarak otozomal dominanttır ve bazı kesimler tarafından en yaygın tek gen hastalığı olarak kabul edilmektedir. Mutasyonun paternal orijini bir çalışmada, 12 aileden 14 ünde bulunmuş (139), geniş ailelerdeki kendiliğinden oluşan mutasyon oranı %50 lerde olduğundan tek-gen etiyolojisi halen kesin olarak saptanmamıştır (284, 287). NF-1, nörofibramatozis vakalarının %90 ını meydana getirmekte ve yaşamın ilk 5 yılında ortaya çıkmaktadır. Deri pigmentasyonu (cafe-au-lait lekeleri) ile karakterize edilmekte, derideki multipl nörofibromlar ve olası daha derin periferal sinirler ve nöral kökleri içermektedir. NF-1 olgularının büyük çoğunluğu (%94), Lisch nodülleri veya pigmentli iris hamartomları (1. doku veya hücrelerin yanlış embriyonal gelişmesi sonucu oluşabilen tümör benzeri lüzumsuz doku kitlesi. 2. dokudaki 24
46 kusurlu gelişime bağlı olarak meydana gelen tümöre benzer yumru şeklinde oluşum, hamartom. 3. Yeni oluşmuş kan damarlarından meydana gelen tümör.) bulundururlar (277). Ayrıca, optik sinir gliomları, astrositomlar, ependimomlar, akustik nöromlar, nörilemma, meningiom ve nörofibroma deneyimleri de olabilir. NF-1 olgularının %4-45 inde beyin tümörleri de gözlenebilir (28, 121, 321). Airewele ve arkadaşları, NF-1 taşıyan bayanların erkeklere oranla, neoplazi geliştirme risklerinin daha fazla olduğunu rapor etmişlerdir (4). Ayrıca etkilenmemiş birinci dereceden akrabalarda, kanser riskinin artış sergilemediğini gözlemlemişlerdir. Bunun sonucu olarak, probanttaki olası bir malignansiye, genel kanser riskinde dikkate değer bir etkisi olan modifiye olmuş genin neden olmadığı anlaşılmıştır (4). Son birkaç yıl içinde, NF-1 e neden olan NF-1 geni tespit edilmiş ve bu gen tarafından kodlanan protein, nörofibromin ayrıntılı araştırma altına alınmıştır. NF-1 hastalarından alınan tümörlerle yapılan çalışmalar, NF-1 geninde homozigot delesyon taşıyanlarda, NF-1 in tümör süpresör rolü oynadığı ortaya konmuştur (348) Nörofibromatozis Tip 2 Nörofibromatozis tip 2 (NF-2), veya bilateral akustik nörofibramatozis, NF-1 in 1/10 i kadar sıklıkta gözlenmektedir. NF-2, NF nin merkezi formu olarak kabul edilmekte, 50,000 kişide 1 gözlenmekte ve tüm NF olgularının %10 unu kapsamaktadır. Otozomal dominant olarak kalıtılan bir hastalıktır. Moleküler biyoloji dilinde, kromozom 22 de defekt bulunmaktadır. Olguların %25 inde meningiomlar NF-2 ile ilişkilidir. NF-2, multipl tümörlerin klinik karakteristiklerini sergilemekte, genellikle de kraniyal ve spinal sinir uçlarının schwannomları dikkat çekmektedir. 25
47 Multipl ependimomlar, kraniyal kavite ve spinal korddaki araknoidal hücrelerden gelişen meningiomlar ve spinal kord veya beyin kök astrositik gliomları NF-2 li bireylerde ortaya çıkmaktadır. Bunlar genellikle düşük-dereceli malignansiler olmaları yanında, yıkıcı nörolojik etkilere sahiptirler (209). Kromozom 22 nin uzun kolundaki bir delesyon ile ortaya çıkan NF-2, meningiomlar, gliomlar ve spinal nörofibromlarla ilişkilidir (302, 359) Yeni Tanımlanan Tümörler Atipik Koroid Pleksus Papillomu (WHO derece II) Koroid pleksus epitelinden köken alan ventrikül içi papiller neoplazilerin çoğu benign tabiata sahiptir ve cerrahi olarak çıkarılması ile kür sağlanabilmektedir. Bu spektrumun öteki tarafında ise koroid pleksus karsinomları yer almaktadır. WHO çalışma grubu tarafından her ikisi arasında özelliklere sahip bir olgu sunulmuştur; atipik koroid pleksus papillomu. Koroid pleksus papillomundan mitotik aktivitede artma olması ile ayrılmaktadır. Muhtemelen cerrahi ile kür mümkündür ancak tekrarlama riski önemli ölçüde yüksektir Ekstraventriküler Nörositoma (WHO derece II) Nörositoma tanımı akla serebral nöroblastomadan farklı patolojik özelliklere sahip nöral bir tümörü getirmektedir. Sıklıkla genç erişkinlerde görülmektedir ve foramen Monroe bölgesi lateral ventrikülde yerleşmektedir. Serebral hemisferlerde gri cevher ya da beyaz cevher etkilenebilmektedir ve tipik bir radyolojik görüntüsü bulunmamaktadır. Sıklıkla iyi sınırlı, kistik tabiatta olabilen değişken kontrastlanma ve peritümöral ödem paterni gösteren tümörlerdir. Genellikle iyi prognoza 26
48 sahiptirler. Son yıllarda yayınlan çalışmalarda ventiküler sistem dışında görülen parenkimal yerleşimde bildirilmiştir. Bu olgularda da benzer biyolojik davranış ve histopatolojik bulgular tarif edilmiştir. Ekstraventriküler nörositom (EVN) nadir bir tümör olmasına karşın görüntüleme özellikleri ile diğer tümörler ile benzerlik gösterebilmektir ve preoperatif dönemde doğru teşhis koymak zor olabilmektedir. Bu nedenle genç bir hastada büyük boyutlu, serebral parenkimal yerleşimli, içerisinde kistik nekroz, kalsifikasyon gibi bulgular gösterebilen, iyi kontrast tutabilen bir kitle lezyonu görüldüğünde EVN nin akla gelmesi önerilmektedir Papiller Glionöronal Tümör (WHO derece I) Papiller glionöronal tümörler geniş bir yaş grubunda görülebilmektedir ve klinik olarak benign tümörlerdir. Özellikle temporal lobda görülürler. BT ve MRG de kontrast tutan, iyi sınırlı kitle lezyonu olarak görülürler ve nadiren kistik-mural nodullü bir patern gösterebilirler. Literatürdeki olguların sadece 3 ünde kortikal gri cevherde yerleşim görülmüş, diğerleri beyaz cevherde yerleşmiştir. Genellikle iyi sınırlıdırlar ve hafif peritümöral ödem gösterirler. Radyolojik olarak kalsifikasyon gösterilebilir. Bazen tümör içi kanama ile başvurabilmektedir ve kavernoz hemangiom ile karışabilmektedir. İkibinaltı yılında yayınlanan 21 olguluk bir çalışmada (67) 3-84 aylık takip sonucunda herhangi bir ek tedavi uygulanılmaksızın tümör rekürrensi olmadığı bildirilmiştir Ventrikülün Rozet Oluşturan Glionöronal Tümörü (WHO derece I) İlk önceleri serebellumun Disembriyoplastik Nöroepitelyal Tümörü olarak tarif edilmiştir. Nadir görülen, yavaş büyüme paternine sahip olan ve 4. ventrikül 27
49 bölgesinde görülen bir tümördür, sıklıkla genç erişkinleri etkiler (ortalama yaş 33). Obstrüktif hidrosefaliye neden olabilir, ataksi en sık klinik bulgusudur. Tipik olarak orta hattan köken almaktadır ve ilk olarak serebellumu ve 4. ventrikülün duvarlarını tutar. Sıklıkla 4. ventrikülü ve akuaductu doldurur, parenkimal uzanım gösterebilir. MR da iyi sınırlı, heterojen, kısmen kistik olabilen tümöral lezyon T1 ağırlıklı MR da hipointens, T2 ağırlıklı sekanlarda hiperintens görünmektedr. Kontrastsız BT de hipodens görülmektedir. Benign klinik davranışları dikkate alındığında muhtemelen cerrahi ile kür sağlanabilecek bir tümördür Pineal Bölgenin Papiller Tümörü (WHO derece II/III) Pineal bölgenin yeni tanımlanan nörepiteliyal tümörü çocukluk ve yetişkin yaş gruplarında görülebilir (ortalama yaş 3). Oldukça büyük (2,5-4 cm) boyutlarında iyi sınırlı bir tümördür. MR da T1 de düşük, T2 de ise artmış intensitede görülür ve kontrast tutmaktadır. Her ne kadar makroskobik olarak pineositomadan ayrılamasa da histolojik bulgular pineal parenkimal tümör ile uyumlu değildir. Pineal bölgenin papiller tümörleri farklı biyolojik davranış gösterebilirler. Buffenoir ve arkadaşları tarafından yayınlan bir olgu sunumu ve literatür derlemesinde, ameliyat sonrası 5 yıllık dönemde yüksek rekürrens riski olduğu bildirilmiş ve cerrahi rezeksiyon derecesinin prognozu etkilediği belirtilmiştir. Bu nedenle tam cerrahi rezesiyonun hedeflenmesi ve radyoterapi uygulanılması önerilmektedir (39). 28
50 Pütisitom (WHO derece I) Pütisitomlar nadir görülen, solid, düşük dereceli, iğsi hücreli glial tümörlerdir. Yetişkin yaş grubunda nörohipofizden ya da infindibulumdan köken almaktadır. Geçmişte sellare parasellar bölgenin diğer (örn granular hücreli tümör ya da pilositik astrositom) tümörleri içinde bu terim kullanılmıştır. Şu anda ise nörohipofizden yada infundibulumdan köken alan düşük dereceli glial tümörler için kullanılmaktadır. Pilositik astrositomlardan farklıdırlar. Klinik olarak görme bozuklukları, baş ağrısı ve hipopituitarism bulgu verirler. Genellikle iyi sınırlı, solid tümörlerdir ve birkaç santimetre boyuta ulaşabilirler. Genel olarak bu tümörler opere edildiklerinden doğal seyirleri tam olarak bilinmemekle beraber Wolfe ve arkadaşları bu tümörün ileri derecede vasküler olabileceğini bildirmişler ve kısmi rezeksiyon ile rekürrens riski olması nedeniyle mümkün olabilen olgularda tam rezeksiyon yapılması gerekliliğini önermektedirler (9, 358) Adenohipofizin İğsi Hücreli Onkositomu (WHO derece II) İğsi hücreli onkositom anterior hipofiz bezinin endokrin olmayan onkositik tümörü olarak tanımlanır (ortalama yaş 56). Makroskobik olarak fonksiyon göstermeyen hipofiz adenomlarından ayrılamazlar ve benign bir klinik seyre sahip oldukları düşünülmektedir (9, 39, 67, 191, 349, 358, 365) Beyin Tümörlerinin Ailesel İlişkileri SSS tümörleri ile kalıtsal sendromlar arasındaki ilişkiye ek olarak, beyin kanseri etiyolojisinin araştırılması, SSS ve diğer kanserleri bir noktaya toplayan SSS tümörlü hastaların ailelerine odaklanmaktadır (85, 175, 177, 178, 179, 180, 331). Bu gibi 29
51 kanser ailelerinde hastaların ve yakınlarının tümörleri, histolojik ve biyolojik olarak benzerdir ve iyi belgelenmiştir, fakat genetik ve SSS neoplazmları arasındaki kesin ilişki halen bilinmemektedir (330). Metodolojik kısıtlamalar, bu çalışmaların birçoğunun otoritesini sınırlandırmaktadır. Ayrıca, ailelerin yaygın olarak neoplazi indüksiyonuna katkıda bulunan çevresel ajanlara maruz kalmaları da bu çalışmaları karmaşık hale getirmekte, fakat kardeşlerdeki benzer beyin ve diğer tümörlerin varlığını (69, 85), ve kanser ailesi sendromunu birbirleriyle tutarlı olarak rapor etmektedirler (177, 178, 179, 180, 197, 198). Etiyoloji bakımından, ailesel vaka ile ilgili iki olası açıklama ortaya çıkmaktadır (4). SSS tümörlerinin ailesel kümelenmesine bir genetik faktör neden olabilir, veya (8) maruz kalmalara kalıtsal bir eğilim, yığılmaları üretebilir Beyin Tümörlerinin Ailesel İlişkileri - Ailesel Yığılma Bir ailedeki hastalığın çok kuşaklı öyküsü, genetiğin yanında, yaygın çevresel maruziyet olasılıklarını işaret etmektedir (30, 114, 204, 362). İkiz çalışmalarının basit bir genetik etiyolojiyi işaret etmesine rağmen, bu agregasyonun yalnızca şans sonucu ortaya çıkmadığı, bir takım faktörlerin sonucu oluştuğu, beyin tümörlü 250 çocukla yapılan bir aile çalışması ile gösterilmiştir (30). Segregasyon analizleri agregasyonun multifaktöriyel kalıtım sonucu ortaya çıktığını işaret etmektedir. Altıyüz erişkin glioma hastalarını içeren aileler ile yapılan başka bir çalışmada, segregasyon analizleri ile oluşturulan poligenik bir modelin beyin tümörlerinin oluşma şekillerini en iyi şekilde açıklamaktadır (64). 30
52 İki yüz doksan yedi glioma ailesinin birinci dereceden yakınını kapsayan segregasyon analizleri, multifaktöriyel bir modeli reddetmemekle birlikte, otozomal resesif bir modelin en uygun olduğunu savunmaktadır (205). Bu çalışma glioma vakalarının %5 inin familiyal olduğunu varsaymaktadır. Grosman ve arkadaşları (101), beyin tümörlerinin ailelerde bilinen kalıtımsal bir hastalık olmadan ortaya çıkabileceklerini ve çok sayıda ailede çevresel nedenler sonucu oluştuklarını göstermişlerdir. Kalıtımsal mutant p53 ile ortaya çıkan kalıtımsal Li-Fraumeni kanseri aile sendromuna sahip ailelerinin keşfinden sonra, p53 ün beyin tümörlerini de kapsayan çok sayıda insan kanserindeki önemini ortaya koyan çalışmalar yapılmıştır (234). Li ve arkadaşları (178) glioma geliştiren erişkinlerle yaptıkları populasyon-bazlı çalışmada, kontrol vakaları ile karşılaştırıldığında, p53 mutant tümörlü olguların birinci-dereceden kanserli yakınları olduğunu ortaya koymuşlardır ve diğer vakalarda daha önceden kanser varlığı ortaya konmuştur. Germ-line p53 mutasyonları sıklıkla multifokal glioma, glioma ve başka bir primer malignansiye sahip olgularda, yada diğer beyin tümörlü olgulara göre ailelerinde kanser hikayesi olduğu ortaya konulmuştur (167). Günümüzde bu alandaki çalışmalar, tümörlerdeki p53 mutasyonu sıklığını belirlemek ve spesifik p53 mutasyonları ile spesifik maruz kalmalar arasındaki korelasyona yoğunlaşmıştır. p16, Rb, ve MDM2 gibi tümörlerdeki önemli diğer hücre döngüsü düzenleyicilerinin değişimleri de değerlendirmiştir. Mutasyon, delesyon ve amplifikasyona uğramış genlerdeki germline mutasyonları belirlemek amacıyla sporadik gliomlarda dizayn edilmiş bir 31
53 çalışmada, CDK4, p16, ve p15 genlerinde germ-line mutasyonlarına yönelik bir kanıt bulunamamıştır (94) Metabolik Yatkınlık ve Polimorfizmler Primer beyin tümörlerinin yalnızca küçük bir yüzdesinde kalıtılan nadir mutasyonlar bir faktör olarak karşımıza çıkmaktadır. Bundan dolayı, araştırıcılar dikkatlerini beyin tümörlerine yatkınlık oluşturan veya daha agresif seyretmelerini sağlayan genlerdeki çevresel faktörlerin de etkileri ile ortaya çıkan yaygın polimorfizmlere çevirmişlerdir. Beyin ve diğer tümörlerin yatkınlığına neden olan genlerdeki değişimler; oksidatif metabolizmayı, karsinogenlerin detoksifikasyonunu, DNA stabilitesini ve tamirini veya immun yanıtı etkileyebilmektedirler (143). Karsinogenik maruz kalınmalar sonucu ortaya çıkan genetik polimorfizmlerin primer olarak sigara tüketimi ile ilişkili olduğu çeşitli çalışmalarla gösterilmiş, fakat genetik teknolojisindeki son gelişmeler diğer kanserlerde de yatkınlık oluşturan polimorfizmlerin epidomiyolojik değerlendirilmesini sağlamıştır (76), ilk kez sitokrom p450 2D6 (CYP2D6) ve glutatyon transferaz tetanın (GSTT1) dikkate değer bir şekilde beyin tümörü riskini arttırdıklarını göstermişlerdir (147). GSTT1 genotipinin, oligodendrogliom oluşumunda artmış risk oluşturduğunu bulmuşlardır (3). Erişkinlerde glioma riski ile GSTM1, GSTT1, ve CYP1A1 null genotipleri arasında istatistiksel olarak anlamlı bir ilişki bulamamışlar fakat; hızlı N-asetiltransferaz asetilasyonu ile neredeyse iki kat artmış risk ve orta asetilasyon için ise %30 artmış risk tespit etmişlerdir. Fakat bu bulgu, gliomlu erişkinlerde yapılan başka bir olgu- 32
54 kontrol çalışmasında desteklenmemiştir (254, 259, 50). Oligoastrositomlu hastalarda, ERCC1 in nükleotidinde AA veya AC ve CC genotipine sahip kontrollere göre 4.6 kat (%95 güven aralığı ) olduğunu göstermişlerdir (50). Fakat bu genotiplerin odds ratioları, glioblastomlu hastalar ile kontrollerde neredeyse aynı olarak bulunmuştur. Bu varyantın sessiz bir mutasyon olmasına rağmen; ERCC1 mrna stabilitesini etkilemekte, ve aynı polimorfizm bir nükleolar proteinde ve T-hücre reseptör kompleksi altünitesinde lizin amino asidinin glutamine değişimine neden olmaktadır. Aynı populasyonlar kullanılarak Caggana ve arkadaşlarının yaptıkları çalışmada, ERCC2 nin AA genotipinin (C-A polimorfizmi [R156R) CC veya CA genotiplerine göre glioblastoma, astrositom veya oligoastrositomlu hastalarda kontrole göre istatistiksel olarak anlamlı bir şekilde daha yaygın olduğunu tespit etmişlerdir (42, 50). Ayrıca bu varyant bir sessiz polimorfizmdir, bu da başka bir genin buna eklendiğini ve tespit edilen ilişkilere neden olabileceğini düşündürmektedir. Bunlara ek olarak; kan testlerinden elde edilen genotip verileri, gliomlar ile ilgili bu populasyon-bazlı çalışmalardaki en düşük sağkalımı sergileyen hastalar için mevcut değildir, bu polimorfizmlerin sağkalım veya etiyoloji ile ilişkili olup olmadığı kesinlik kazanmamıştır. Bu verilerin teyit edilmesi için daha kapsamlı çalışmaların yapılması gereklidir. Daha geniş hasta grubu ile yapılacak olan çalışmalar, örnek sayısının azlığından kaynaklanacak olan yanlış pozitif sonuçların veya var olan bağlantılarının tespit edilememesi gibi problemleri ortadan kaldıracaktır. 33
55 Tümör Süpresör Genler p53 p53 mutasyonu tüm astrositk tümörlerde gözlenmesine rağmen anaplastik astrositomda (%64) glioblastoma multiformeden (%26) daha fazla bulunmuştur (146). Bu, p53 ün p16 ve p21 gibi tümör inhibitör proteinlerle harmonize çalışmasını engelleyerek, daha malin formlara dönüşümünü açıklamaktadır. p53 genindeki mutasyon sekonder glioblastomda %65 iken, primer glioblastomda sadece %10 olarak bulunmuştur (351). p53 proteinin aşırı ekspresyonu p53 geninin mutasyonunu yansıtmaktadır. p53 proteininin aşırı ekspresyonu sekonder glioblastomlarda %90 dan fazla iken, primer glioblastomlarda %35 ten az bulunmaktadır (351). Düşük dereceli astrositomlarda görülen en sık görülem kromozomal değişiklik kromozom 17p13.1 delesyonu ve bu bölgede yerleşim gösteren tümör süpresör gen olan p53 (TP53) mutasyonlarıdır (303). Buna ek olarak germline p53 mutasyonlarını taşıyan Li-Fraumeni sendromlu hastalar, artmış gliom riski taşımaktadırlar. Bu da p53 değişimlerinin bazı astrositomların başlangıcına dahil olma hipotezini desteklemektedir. DNA hasarının farklı derecelerine yanıt olarak, TP53, proapoptotik genleri ve hücre döngüsü inhibitörlerini transaktive etmektedir (250, 381). Bu nedenle, normal p53 taşımayan hücreler hücre döngüsü aresti ve apoptoz yeteneklerini kaybederler. p53 mutasyonları düşük dereceli astrositomların yaklaşık %65 inde gözlenmesine rağmen, benzer bir spektrumun p53 mutasyonları, anaplastik astrositomlu ve glioblastomlu olguların alt kümelerinde bulunmakta ve 34
56 bu durum da anaplastik astrositomların ve bazı gliomların daha erken safhadaki bir düşük dereceli astrositomdan geliştiğini işaret etmektedir. Sporadik düşük dereceli astrositomun oluşumunun başlangıcında, p53 mutasyonunun rolü çok iyi anlaşılabilmiş değildir. Çünkü kısmen, diğer kanserlerin tersine preneoplastik olarak varsayılan lezyonlarda (örneğin: PIN prostat kanseri ile ilişkilidir), hiçbir preneoplastik lezyon gliomlar ile ilişkili değildir. Buna rağmen, germline p53 mutasyonlu bir hastanın tümörlerinde p53 değişimleri dikkatlice değerlendirildiğinde, Fulci ve arkadaşları astrositik tümölerin klinik olarak tespit edilemeyen prekürsörleri için genetik bir kanıt bulabilmişlerdir (92). Bu önemli genetik bulgular, preastrositom lezyonlarının varolabileceğini ve nadir rastlanan p53 somatik mutasyonları sonucu oluşabileceklerini göstermektedirler. Sekonder glioblastomların, p53 mutasyonu gibi diğer anomalilerle birlikte, düşük dereceli ve astrositomlarda gözlemlenen birçok değişikliğe sahip olduğu bulunmuştur. Tümör süpresör gen olan p53, apoptozun kontrolü, p21 ve siklin bağımlı kinazların aracılığı ile hücre döngüsünün regülasyonu, büyüme arresti, hücre siklusunun inhibisyonu, genotoksik ya da hücresel strese yanıt olarak farklılaşma, tümör anjiyogenezinin kontrolü ve hızlandırılmış DNA tamiri ya da yaşlanmada yer alan birçok genin regülasyonu gibi birçok önemli görevde yer almaktadır (37). p53 geni 17. kromozomun kısa kolunda 13,1. bant üzerinde lokalizedir. On bir ekzondan meydana gelir. Transle edilen protein 393 aminoasit (aa) içermektedir. Protein 3 ana domain içermektedir: transaktivasyon, aa 1-42; DNA bağlaması, aa ; oligomerizasyon, aa Protein tetramer olarak organize olmuştur. Ultraviyole ışık, karsinojenler ve radyasyon normal timidin fosforilazı aktive edebilir. 35
57 Bir onkogen olan MDM-2 p53 ün amino terminal kısmına bağlanır ve p53 ün down-regülasyonu na neden olur (37). p53 te meydana gelen mutasyonlar genomik kararsızlık ve kansere karşı artmış duyarlılıkla ilişkilidir. Tüm kanserler arasında en sık mutasyona uğrayan proteindir. Serviks tümörlerinde %90, kolorektal tümörlerde %70 oranda insidans gösteren birçok tümörde p53 mutasyonu bulunmaktadır. p53 mutasyonu, Li- Fraumeni sendromu gibi belirli ailesel kanserlerin daha yüksek insidans ile sonuçlanan kalıtımsal mutasyonları içeren birçok yolakta inaktive olabilir. İnsan adenovirüsü ve papilloma virüsleri gibi belirli DNA tümör virüsleri proteine bağlanırlar ve inaktive ederler. Fonksiyonel p53 ün tümörogenez üzerinde koruyucu bir etkisi olduğu düşünülmektedir (106, 99). İlaç resistansının geliştirilmesinde yer alan DNA tamir yolaklarının varlığı, son yıllarda tümör hücre hatları üzerine yapılan in vitro çalışmalarda giderek artan bir şekilde gösterilmektedir. Tümör örneklerinde, DNA tamir yolaklarında yer alan spesifik genlerin ekspresyonlarının ölçümü DNA tamirinin olası klinik önemini değerlendirmek için kullanılmaktadır. Tümörlerde p53 proteinin yükseltilmiş seviyeleri p53 genindeki mutasyonu işaret etmektedir. Hücre döngüsü kontrol noktalarının regülasyonunda yer alan p53 proteini gibi, gende bulunan DNA tamir ve apoptotik yolakların mutasyonları tümör hücrelerinin sitotoksik ilaçlar üzerindeki duyarlılığındaki değişiklikten sorumlu olabilir. Bu durum, ilaç direnci ile sonuçlanabilir. Artmış p53 seviyelerinin immunohistokimyasal olarak saptanması, over, meme ve mesane kanserlerini içeren birkaç tümör tipinde agresif fenotipin ve zayıf prognozun özellikleriyle ilişkilidir (38). 36
58 p53, genomik hasarın ve hücresel stresin birçok türüne karşı oluşturulan hücresel yanıtı regüle eden önemli bir ağın merkezi düğüm noktası olarak düşünülmektedir. Bu durum, p53 te meydana gelen defektlerin insan kanserlerinde neden bu kadar yaygın olduğunu açıklamaktadır. Fonksiyonel p53 genine sahip olmayan kanserler, DNA zincir kırıklarını ve anaploidiyi içeren daha fazla DNA hasarını tolere edebilecekler, uygun olmayan proliferasyon sinyallerine ve nükleotid stresine göz yumacaklar ve senesense daha zor gireceklerdir (300) RB1 (Retinoblastoma) Astrositomda onkogenezi başlatan ve daha malin formlara dönüşümden sorumlu genlerden biri retinoblastoma (Rb1) tümör baskılayıcı genidir. Retinoblastom geni 13q14 bölgesinde yerleşmiştir. Normalde, yabanıl Rb geninin kodladığı Rb proteinleri fosforlanarak (hipofosfarilasyon) hücre içi E2F proteinini (pe2f) bağlar ve G1 kontrol noktasında siklusu durdurur. Retinoblastoma geninin yokluğu ve mutant olması durumunda, ya Rb proteini fonksiyon görmeyecek veya Rb1 proteininin uygunsuz fosforlanması gerçekleşecektir. Bu durumda, Rb1 ile E2F proteinleri birbirine bağlanamayacağından, hücrenin S fazına geçmesi gerekli E2F proteinleri gibi transkripsiyonel faktörlerin artmasına neden olur (40, 68, 137, 377). Bu bölgedeki heterozigosite kaybı, düşük dereceli astrositomada yaklaşık %20, anaplastik astrositomda yaklaşık %25 ve glioblastomda yaklaşık %35 bulunmuştur. Ancak Rb geninin mutasyonu çok düşük oranlarda bulunmuştur (377). Kromozom 13q bölgesindeki heterozigosite kayıpları primer glioblastomlarda düşük düzeylerde gözlenirken (%12), sekonder glioblastomlarda daha yüksek düzeyde gözlenmesi 37
59 (%38), Rb1 geninin de gliomaların malin formlara dönüşümünde olaya katıldığını göstermektedir (231). Genetik olarak, modifiye edilmiş normal insan astrositlerini kullanarak Sonoda ve arkadaşları (320) anaplastik astrositoma gelişiminde kritik olan yolakları tanımlamayı başarmışlardır. İnsan telomeraz katalitik komponentinin ekspresyonu, Ras yolağı aktivasyonunun (233), ve p53 RB1 in kombine inaktivasyonu gibi genetik değişiklik normal astrositlerini anaplastik astrositomlara (WHO derece 3) transforme edebilmektedir. İlginç olarak; bu tümörler, yalnızca Ras yolağı aktivasyonu mutant H-Ras ile indüklendiğinde gelişebilmektedirler. Bunun aksine, fosfotidil inositol 3- kinaz yolağı (Akt ile başlatılan) veya EGFR aracılığı ile Ras aktivasyonu, anaplastik astrositom oluşumunu indükleyememektedirler. Düşük dereceli astrositomlarda, belki de hücreler yetersiz olarak farede tümörojenik olmak için transforme olduklarından dolayı gözlenememektedirler. Buna karşın; bu araştırmalar anaplastik astrositomların sınırlı sayıda spesifik primer genetik değişimler sonucunda oluştuğunu işaret etmektedir. Bunlara ek olarak, anaplastik astrositom oluşumunda Akt veya EGFR aracılıklı Ras yolağı aktivasyonu ve insan telomeraz katalitik komponenti/p53/rb1 arasındaki kooperasyon eksikliği, primer glioblastom oluşumuna göre bu tümörlerin oluşumunda etkili olan moleküler olayların farklı olabileceğini düşündürmektedir PTEN (Fosfataz ve tensin homoloğu, MMAC 1) PTEN geni kromozom 10q23.3 bölgesinde yerleşmiş tümör baskılayıcı gendir ve iki araştırıcı tarafından 1997 yılında saptanmıştır (181, 322). Bu nedenle PTEN/ 38
60 MMAC1 olarak bilinir ve hücrelerin farklılaşma ve sağkalımında etkilidir. PTEN lokusunun kaybı gliblastomlarda %80 dolayında bildirilmesine rağmen PTEN mutasyonu %20 30 dolayında ve daha yaşlı hastalarda rapor edilmiştir (31, 269). Ancak PTEN geni mutasyonu primer glioblastomda %32, sekonder glioblastomda %4 dolayında bulunmuştur (19, 276, 303, 332, 381). Diğer yandan primer glioblastomda 10. kromozom un q ve p kolunu içeren tüm bir allelinin kaybına karşın, sekonder glioblastomda LOH lar, kısmi delesyonlarla sadece q kolunda bulunmuştur (90, 91). Tümör evolüsyonunda 10q da olan LOH ve PTEN mutasyonunun, p53 deki değişiklikleri takiben tabloya eklendiği kabul edilmektedir. P53 mutasyonu gösteren glioblastomlarda PTEN lokusu dışında özellikle 10q25-qter bölgelerinde kayıplar saptanmıştır ki bu sekonder glioblastom gelişiminde başka tümör baskılayıcı genlerin varlığını düşündürmektedir (90). PTEN geni 10q23.3 bölgesinde bulunan dual-spesifik fosfatazdır ve sitoiskelet proteini olan tensine homoloji gösterir (303, 381). PTEN, antagonize edici fosfotidilinositol-3,4,5 trifosfat kinaz aracılığı ile fonksiyon gösterir (155). PTEN, fosfotidilinositol-3,4,5 trifosfat yolağının aktivasyonuna neden olur. Bu ailenin bir üyesi Akt dir. PTEN in gliomagenezdeki fonksiyon mekanizması ve rolü birçok çalışmada kapsamlı olarak çalışılmıştır (34,155, 276). Önemli bir şekilde, yapılmakta olan çalışmalardan elde edilen bilgiler, PTEN de meydana gelen mutasyonun, yüksek dereceli gliom görülen olgularda zayıf sağkalımla ilişkili olduğunu işaret etmektedir (285, 314). Tümör süpresör fonksiyonunun yanı sıra, PTEN geni tarafından kodlanan protein, glioblastomlarda gözlenen, anjiyogenez, migrasyon ve invazyonu içeren hücresel agresifliğin de rol oynaması muhtemel bir çok sürecin 39
61 regülasyonunu sağlamaktadır (82,142, 162, 163,164, 248, 288, 339). Bu nedenle, PTEN yüksek dereceli gliomlar için teröpatik bir hedef olarak araştırılmaktadır. Diğer tümör süpresör genler gibi, PTEN hücre proliferasyonunu baskılamaktadır ve apoptozu arttırmaktadır (155). Buna ek olarak, son bulgular, PTEN nin aynı zamanda hücre göçünü de regüle ettiğini göstermektedirler. Örneğin, PTEN, hücre migrasyonunun önemli bir regülatörü olan FAK ın (fokal adezyon kinazı) (141) hücresel hedefidir. Önemli bir şekilde, PTEN kaybı gliom invazyonunda önemli olan matriks metalloproteazı 2 ve 9 u upregüle eder (163, 248). Glioblastomlarda PTEN yolağının hedeflenmesi hücresel proliferasyonu önler, apoptozu arttırır ve invasyonu azaltır. PTEN yolağının inhibisyonundaki en çok teröpatik potansiyele sahip olan küçük molekül bir rapamsin analoğu olan CCI 779 (19, 102). Hem rapamisin hem de CCI 779, tümörlerde aktive edilmiş bir protein kinaz olan mtor u (rapamisinin memelilerdeki hedefi) inhibe eder (307). Rapamisin ve CCI-779, nonglial tümörlerde önemli aktivite gösterir (373, 257) ve gliomlardaki kullanımı günümüzde önemli bir araştırma alanıdır. Kromozom 10 da gerçekleşen heterozigosite kayıpları glioblastomlarda en sık karşılaşılan sitogenetik değişikliklerden biridir ve olguların %80 inde vardır. Bunlar en sık 10p, 10q23 ve 10q25-26 bölgesinde olur (131, 144, 268). Kromozom 10 da gözlenen bu kayıplar primer ve sekonder glioblastomlarda eşit sıklıkta gözlenmiştir (91). Ancak 10. kromozomun bir alelinin kaybı (monozomi) daha çok primer glioblastomlarda rapor edilmiştir (91). Kayıplar düşük dereceli astrositomlarda nadiren saptanırken anaplastik astrositomlarda % 40 düzeyinde bulunmuştur (131, 268). 40
62 MGMT (O 6-metilguanin-DNA metiltransferaz) Alkilleyici ajanlar ultraviyole radyasyona benzeyen bir DNA hasarına (genellikle DNA daki O6-G (Guanin), O4-T (Timin) ve O2-T pozisyonları) neden olurlar. Bu ajanlar tarafından değiştirilen bazlar öncelikle pürinlerdir ve oluşan ürün spektrumu kullanılan ajana bağlı olarak çeşitlilik göstermektedir. Memeli hücrelerinde bu gibi yanlış kodlanan alkilasyon lezyonlarını düzeltmek amacıyla DNA tamir mekanizmaları bulunmaktadır. Bu ürünlerin en mutajenik olanı, modifiye edilen zincir duplike olduğu zaman en yüksek Timin ile eşleşme olasılığı olan O6-MeG (O6- metilguanin) dir. O6-MeG, reaktif hücresel katabolitler tarafından küçük miktarlarda endojen olarak üretilir. Memeli hücrelerinde, alkilasyonun ve oksidatif DNA hasarının tamiri için kullanılan iki temel yolak bulunmaktadır. Alkilasyon DNA hasarının onarımı için en direkt mekanizma, DNA tamir proteini olan ve DNA daki alkil gruplarını uzaklaştıran MGMT yi içermektedir (O6-Metilguanin-DNA metiltransferaz ya da AGT). MGMT, O6-MeG (O6-metilguanin) den metil ya da etil grubunu transfer ederek hasarı tamir eder. Bu nedenle, MGMT (foto reaktivasyon gibi) hasarlı bazları uzaklaştırmak yerine onları doğrudan değiştirir. Çünkü bu reaksiyon proteinin tersinir olmayan inaktivasyonu ile sonuçlandığı için, MGMT intihar eden enzim olarak tanımlanmaktadır. Eğer eşleşmemiş olarak bırakılırsa, O6-MeG bileşimi, hem mutagenez hem de karsinogenez ile ilişkili olan G:C A:T geçişine neden olan timin ile eşleşmeyebilir (229, 333). 41
63 O6-Metilguanin-DNA metiltransferaz (MGMT), alkilleyici ajanlar tarafından oluşturulan DNA hasarının tamirinde önemli bir rol oynar. Guaninde O 6 - pozisyonundan alkil gruplarının uzaklaştırılmasıyla, MGMT, GC nin A:T ye transisyon mutasyonlarını engelleyebilir. Bu mutasyonlar, beyin tümörlerinde çoğunlukla TP53 mutasyonlarını içeren varyasyon tipidir. Tümör ilişkili genlerin CpG adalarının promotör hipermetilasyonu, bu genlerin transkripsiyonel inaktivasyonlarına neden olabilir ve bu epigenetik mekanizma, sinir sistemini etkileyenleri de içeren bazı kanserlerde MGMT susturulmasına katılması için gösterilmiştir. Bundan dolayı, genetik ve epigenetik anomaliler arasındaki bağlantı, bu neoplazmlarda bulunabilir. Sonuç olarak, tipik olmayan metilasyon, örneklerin %38 inde oluşmuştur ve astrositik, oligodendroglial ve ependimal farklılaşmaları içeren glioblastomlar, anaplastik gliomlar gibi daha malin olanlarda %50 den yüksek değerler gözlenmiştir. Buna karşın, non-glial tümörler, bu grup nöroblastom, medulloblastom ve beyin metastazı gibi oldukça yüksek malin tümörler içerse bile, %26 sında tipik olmayan MGMT promotör metilasyonu gösterir. Sonuç olarak, TP53 mutasyonları, metilenmiş MGMT tümörlerinin %25 inde (45/180) bulunurken metillenmemiş MGMT tümörlerinin sadece %10 u (30/289) p53 değişiklikleri gösterir (p<0.001). G:C nin A:T ye değişimi metilenmiş tümörlerin %9 undaki CpG bölgelerinde ve metillenmemiş örneklerin %7 sinde oluşur (21). 42
64 RUNX3 (Runt ile ilişkili transkripsiyon faktörü 3) RUNX3 çeşitli karsinomlarda bilinen bir tümör süpresör gendir. RUNX3 ekspresyonundaki aberasyon henüz kutanöz melanomlarda tanımlanmamıştır. RUNX ailesinin 3 üyesi, RUNX1, RUNX2 ve RUNX3 transkripsiyon faktörleri, çeşitli insan kanserlerinin başlangıcı ve progresyonunda önemli olan gelişimsel regülatörler olarak bilinirler. RUNX3 transkripsiyon faktörüdür ve doku farklılaşmasında sıklıkla bağ kuran faktörler ile ilişki ve iskele görevi gördüğü bilinir. RUNX3 proteinleri, nukleusta lokalizedir ve fonksiyonunun aşağı regülasyonunun vasıtası ile çeşitli kanserlerle bağlantılıdır. Çalışmalar RUNX3 proteinlerinin kromatin yeniden modelleme enzimleri ile birbirlerini etkileyerek gen ekspresyonunu regüle ettiklerini göstermiştir. RUNX3 ün özellikle gastrik tümör progresyonunda bulunduğu gösterilmiştir. Bu gen gastrik ve diğer kanserlerde tümör baskılayıcı rol oynar. RUNX3 promotör bölgesinin hipermetilasyonu ekspresyonunu down regüle eder. RUNX3 kromozom 1p36 da bulunur (151) DMBT1 (Malin Beyin Tümörlerinde Silinmiş 1 Geni) Sisteinden zengin çöpçü reseptör (SRCR) süper ailesinin bir üyesi olan DMBT1, bir medullablastom hücre hattında 10q de homozigot bir delesyon olarak tanımlanmıştır. DMBT1 in intragenik delesyonları glioblastomların %25 inde ve birçok beyin tümörü hücre hattında tanımlanmıştır. SRCR süper ailesinin üyeleri immün sistemde ya da diğer dokularda proliferasyonun veya diferansiyasyonun başlaması ve epitelyum hücrelerinin polaritesinin ekstraselüler matriks ve hücre 43
65 yüzey proteinleri arasındaki ilişki aracılığı ile durdurulması gibi farklı fonksiyonlara bağlıdır (223). DMBT1 in genomik yapısı birçok yayında bulunmaktadır. Komşu DMBT1 lokusu mikrosatellit markırlarının alelik kayıpları, glioblastom gibi birçok kanserde tanımlanmıştır. Homozigot delesyon gastrointestinal kanser, akciğer kanseri, oligodendrogliom ve medullablastomada da bulunmuştur. Bu genin fonksiyonu henüz çok net olmamasına rağmen, DMBT1 lokusundaki yüksek frekanstaki delesyonu, kromozom 10 daki olası aday tümör süpresör genlerden biri yapmaktadır (227) PIKE (Fosfotidilinositol 3-kinaz arttırıcı, CENTG1) Fosfotidilinositol 3-kinaz arttırıcı (PIKE) anti apoptotik işlevi olan bir GTP bağlama proteinidir. PIKE ailesinin tek bir genden köken alan (CENTG1) ve alternatif bölünme ya da farklı transkripsiyon başlama bölgesi olan PIKE-L, PIKE-S ve PIKE-A adlarında 3 üyesi vardır. Her iki PIKE-S ve PIKE-L, fosfoinosit- 3 kinaza bağlanmakta ve onun aktivitesini arttırmaktadır. PIKE-A, PI3K ile etkileşim göstermez. Onun yerine yukarı akış efektörü olan Akt ve onun aktivitesini kontrol etmesi ile ilişkilidir. Bu olaylar GTPaz aktivitesi ile ilişkilidir. Çünkü PI3K ve Akt nin her ikisi hücre büyümesini tetikleyen ve apoptoza karşı rol alan büyüme faktörü aracılı sinyalleşmesinin önemli efektörleridir ve bu nedenle PIKE lar büyüme faktörleri ile fizyolojik fonksiyonları etkinleştirmede moleküler anahtar olarak çok önemlidir (48). PIKE, PI 3-kinaz (PI3K) aktivitesini arttıran bir nükleer GTPaz dır. Sinir büyüme faktörü (NGF) muamelesi, PIKE aktivasyonunu PLC-g1 in nükleer 44
66 lokalizasyonunu tetikleyerek gerçekleştirir ki bu da; PIKE için fizyolojik guanin nükleotit faktör olarak rol oynar. PI3K geniş çaplı hücre tiplerinin nükleusunda oluşur ve çeşitli uyarıcılar PI3K nükleer transkripsiyon faktörünü ortaya çıkarır. Sitoplazmik PI3K iyi karakterize edilmiştir. Fakat nükleer PI3K nın biyolojik fonksiyonu hakkında bilgilerimiz azdır. Son zamanlarda; nükleer PI(3,4,5)P3 reseptörü, nükleofosmin/b23 NGFuygulanmış PC12 nükleer ekstraktlarından tanımlanmıştır. PI(3,4,5)P3/B23 kompleksi, kaspaz-aktive edici Dnaz (CAD) ın aktivitesi ile DNA fragmentasyonunu inhibe ederek NGF nin anti apoptotik etkisine aracılık eder. Böylece, PI(3,4,5)P3/B23 kompleksi ve nükleer Akt efektörleri, PIKE/nükleer PI3K sinyalleşmesine NGF ile destekleyerek düzenli bir şekilde aracılık edebilir (367). Sitoplazmik PI3K aktivasyonu, aktive edilmiş reseptör tirozin kinazlar (PDGFR, EGFR, CD28 gibi) ya da Ras gibi GTPaz proteinleri gerektirmektedir. Fakat, bunlardan hiçbirinin PI3K aktivatörü olarak nükleusta bulunduğu bilinmemektedir. NGF li hücrelerin stimilasyonu yine de sitoplazmik benzerinden geçici olarak farklı olan nükleer PI3K yı aktive eder ve sonuçta 3- fosforile olmuş fosfoinositid lipidlerin akümülasyonunu yönetir, bu da nükleer PI3K nın bilinmeyen dengeleyiciler tarafından regüle edildiğini önerir. Aday nükleer GTPaz PIKE etkileşimleri, PI3K ile onun lipit kinaz aktivitesini uyarır. Lizin 413 ve serin 414 rezidülerine GTP bağlayan dominant negatif PIKE, sırasıyla alanin ve asparajinaz değiştirmiş, mutasyona uğramış, nükleer PI3K aktivitesinin NGF arttırmasını önler. PIKE-S GTPaz ve PI3K arasındaki etkileşim, Ras 45
67 ve PI3K arasındaki GTP bağlı etkileşim gibidir. Reseptör tirozin kinazlar ve Ras gibi sitoplazmik PI3K aktivatörleri, iki PI3K alt ünitelerinden birini bağlar fakat, PIKE-S direkt olarak hem p85 hem de p110 ile etkileşir ve bağlanır. PIKE 753 amino asitlik bir polipeptidi kodlar ve moleküler ağırlığı yaklaşık olarak 90 kda olan nukleusta lokalize beyine özgü bir proteindir. PIKE 3 prolinden zengin bir bölüme, bir GTPaz bölümüne ve bir de PH bölümüne sahiptir. PC12 hücrelerinde, NGF muamelesi PIKE etkileşimini başlatır. PIKE-S aktivasyonuna benzeyen zaman akışıyla NGF muamelesini takiben PI3K nükleusta lokalizedir. Fakat PIKE ile nükleer PI3K nın aktivasyonu nükleusa transloke 4.1N, NGF uyarılması tarafından inhibe edilmektedir. Şaşırtıcı bir şekilde, bu etki, onun lipaz katalitik aktivitesinden bağımsızdır (368) Onkogenler EGFR (Epidermal Büyüme Faktörü Reseptörü) Epidermal büyüme faktörü (EGF) ve Epidermal büyüme faktörü reseptörü (EGFR) sinyal yolu; de novo glioblastom gelişimine katılan temel genetik değişikliklerden biridir. EGF geni 4. kromozomda bulunurken, EGFR geni 7q12 de yerleşmiştir. EGFR leri aynı PDGFR de olduğu gibi Thirozin kinaz reseptör ailesine bağlıdır (189, 190). Kromozom 7q12 de lokalize EGFR geninin amplifikasyonu ve proteininin aşırı ekspresyonu gliobastomların yaklaşık %35 inde bulunmuştur (184). Bu genin aktivasyonunun hücre zarının dış yüzeyindeki bağlarında yapısal değişiklik yapması bile hücrede otokrin stimülasyonu başlatması için yeterli olmaktadır. EGFR genindeki yeniden düzenlenmeler fazla protein eskspresyonu ile uyumludur (347). 46
68 EGFR amplifikasyonu gösteren olguların tipik olarak kromozom 10 da kayıpları olduğu saptanmıştır (238). Seçilmemiş glioblastomlarda EGFR amplifikasyonu %30-40 düzeyinde bulunmuştur (190, 238). EGFR amplifikasyonu primer glioblastomda %39-73 iken, sekonder glioblastomada %0-9.5 düzeyinde bulunmuştur (34, 276, 303, 332, 371, 381). EGFR lerinin aktivasyonu mitojen ile aktive olmuş protein kinaz (MAPK) gibi hücrelerin çoğalması ve survisinden sorumlu sinyal yollarının aktivasyonuna neden olmaktadır. MAPK proteinlerinin aşırı ekspresyonu gösteren hastalar, radyoterapiye karşı dirençli ve kötü prognozla uyumlu bulunmuştur (210, 252). Öte yandan MAPK ve AKT aktivasyonu, anaplastik astrositomların glioblastoma dönüşümünden sorumlu tutulmaktadır (220). Üstelik, aynı zamanda EGFR amplifikasyonu da olguların yaklaşık olarak %80 i gen düzenlenmesi ve genellikle aktive edilmiş EGFR ile sonuçlanan mutasyonları göstermektedirler (77). Gliom oluşumunda EGFR sinyalinin rolü ile ilgili mevcut literatürler incelenmiştir (34, 214, 276, 303, 381). Ancak; gliom oluşumundaki rolünün yanı sıra, EGFR nin yüksek dereceli gliomların fenotipiyle ilişkili olabilecek birçok hücresel prosesi regüle ettiğini de vurgulamak oldukça önemlidir (22, 47, 169, 251) Sonuç olarak, EGFR glioblastom görülen olgular için önemli bir teröpatik hedef olarak düşünülmektedir. Sporadik gliomlarda gözlemlenen birçok genetik değişikliğe rağmen, bu değişikliklerin gliom gelişimiyle nedensel ilişkisi hayvan gliom modelleri geliştirilmeden oluşturulamamıştır. Örneğin; fare modelini kullanarak yapılan iki yayın, INK4A/ARF kaybının glioblastomların çok daha hızlı geliştiği hipotezini desteklemektedir (15, 342). Bachoo ve arkadaşlarının yaptığı bir çalışmada (15), 47
69 INK4A ve ARF nin kombine kaybı (ancak, p53, INK4 ya da ARF nin tek başına değil) EGFR aktivasyonuna yanıtta astrosit farklılaşmasına olanak vermiştir. Buna ek olarak, aktive edilmiş EGFR ile transfer edilmiş INNK4A/ARF nöral kök hücreleri ya da astrositlerinin implantasyonu yüksek dereceli astrostik tümörle sonuçlanmıştır. Uhrbom ve Holland (343), kombine K-Ras ve Akt sinyalinin GFAP-pozitif astrositlerden değil ama nestin-pozitif nöroprogenitör hücrelerden glioblastom benzeri tümör oluşturabileceğini göstermişlerdir. Eric Holland ın grubunun (342) yaptığı en son çalışmalardan birinde, INK4A/ARF eksikliği hem netsin-pozitif hem de GFAP-pozitif nöral hücrelerden oluşan glioblastomlarla sonuçlanmıştır. INK4A/ARF kombinasyonu ile birlikte K-Ras aktivasyonu, yüksek dereceli astrositom gelişimi için gereklidir. Bu bulgular, INK4A/ARF lokusunun inaktivasyonunun astrositleri transformasyona karşı duyarlı hale getirdiğini ileri sürmektedir. EGFR ya da K-Ras aktivasyonu gibi sadece birkaç genetik değişikliğin yüksek dereceli, primer astrositik lezyonların gelişimi için yeterlidir. Önemli bir şekilde, EGFR aktivasyonu tek başına bu fare modellerinde tümör oluşumu ile sonuçlanmaz (343). Epidermal büyüme faktörü reseptörü-aracılı sinyal gliom hücrelerinde invasiv fenotipi regüle etmektedir (47, 53, 141, 169, 251) ve mutant EGFR (EGFRvIII), antiapoptotik bir protein olan Bcl-X L yi upregüle ederek gliom hücrelerinde apoptozu baskılamaktadır (46). Buna ek olarak, EGFR primer glioblastom hücrelerinde radyasyon ve kemoterapiye olan rezistansa aracılık etmektedir (169, 151). Bu nedenle, EGFR-aracılı sinyali hedeflemenin insan gliomlarının gelecekteki tedavisi için ümit veren bir potansiyele sahip olması şaşırtıcı değildir. Mevcut ajanlar 48
70 tirozin kinaz inhibitörlerini, anti-egfrviii ve anti-egfr kimerik antikorlarını, antisense oligonükleotitleri ve diğerlerini içermektedir (19, 80, 154, 219, 228) PIK3CA (Fosfoinosit 3 Kinaz, Katalitik, Alfa Polipeptit) Phosphoinositol-3-kinase (Pi3k)/Akt yolağı glioblastomlarda merkezi bir rol oynar. Bu tümölerdeki Pi3k/Akt yolağı aktivasyonu farklı genetik alterasyonlar, PTEN mutasyonu, EGFR gen amplifikasyonu ve yeniden düzenlenmesi ve CTMP promotör hipermetilasyonu sonucu olabilir. PI3K nın p110 alfa katalitik altüntesini kodlayan PIK3CA geni 3q26.3 lokalizedir ve malin gliomaları da içeren farklı insan kanserlerinde aktive nokta mutasyonlarını taşıdığı rapor edilmiştir. Bir çalışma beyin tümörlerindeki PIK3CA mutasyonlarına fokuslanmıştır ve anaplastik oligodendrogliomlar, anaplastik astrositomlar, glioblastomlar ve medullablastomalarda aberasyonlar saptamıştır. Diğer çalışmacılar 12q14teki CDK4 e yakın bulunan PIKE nin (CENTG1 olarakta bilinir), glioblastomda sıklıkla amplifiye olduğunu rapor etmişlerdir. PIKE amplifikasyonunun fosforile Akt ye bağlanan ve onun aktivitesini artıran ve böylece hücre göçüne ve apoptozun inhibisyonuna neden olan PIKE-A transkriptinin ekspresyonunu arttırdığı şeklinde sonuçlandığı bulunmuştur (290). Bir çalışma beyin tümörlerinde PIK3CA mutasyonları üzerine odaklanmış ve anaplastik oligodendrogliomlarda, anaplastik astrositomlarda, glioblastomlar ve medullablastomlarda aberasyonlar saptamıştır (36). PI3K/Akt sinyalleşmesinin anormal aktivasyonu insan glioblastomlarının çoğunda bulunmuştur ve hücre döngüsü kontrolü, apoptoza direnç, artmış hücre 49
71 göçü ve invazyon ve anormal neoanjiyogenez gibi çeşitli biyolojik özelliklere katkıda bulunmuştur (119, 155, 52). Yüksek PI3K/Akt sinyalleşmesi, farklı genlerde genetik veya epigenetik alterasyonlar sonucu olabilir; EGFR amplifikasyonu, PTEN mutasyonu ve CTMP hipermetilasyonu glioblastomlarda saptanan en yaygın değişikliklerdir (36, 157, 290). PIKE (fosfotidil-3-kinaz zenginleştirici), Akt aktivitesini uyaran ve hücresel invazyona aracılık eden bir GTPaz dır (3). Over ve serviks karsinomları gibi farklı epitelyum hücrelerinde amplifiye olan PIK3CA, bir proto onkogen olarak rapor edilmiştir (199, 304). Aktive PIK3CA mutasyonlarını 4/15 i (%27) glioblastom olan çeşitli tümörlerde saptandı (290). Fakat 105 glioblastomlu olguyla çalışılan başka bir çalışmada PIK3CA mutasyon oranı sadece %5 olarak saptanmıştır (36). Tüm glioblastomlar PIK3CA gen amplifikasyonu ve aşırı ifadesi gösteren (156) aktive PIK3CA alterasyonları az sayıda glioblastomda saptanmış ve Broderick ve arkadaşlarının çalışmalarındaki PIK3CA mutant glioblastomlu olgulardan hiçbiri bir PTEN mutasyonu taşımamaktaydı (36). Böylece, PIK3CA mutant tümörlerinin küçük fraksiyonunda her aberasyon dışlanmıştır. 12q14 deki CDK4 geni glioblastomların %10-15 inde, yakın genler ile örneğin SAS, GLI1 ve/veya MDM2 ile amplifiyedir (273). Ahn ve arkadaşları, CDK4 e yakın olan PIKE (CENTG1) nin glioblastomlarda genellikle CDK4 ile ko-amplifiye olduğunu rapor etmişlerdir. PIKE amplifikasyonu PIKE-A nın artan ekspresyonu ile ilişkilidir. PIKE-A nın transloke ürünü aktife Akt ye bağlanır ve Akt kinaz aktivitesini uyarmaktadır (3). Glioblastomlarda PIKE ve CDK4 değişmez ko-amplifikasyonunu olguların %12 sinde saptanmıştır. Ayrıca PIKE amplifiye glioblastomlarda PIKE-A 50
72 mrna seviyesi artmaktadır. PIKE-A geninin aşırı ifadesinin tümörlerde amplifikasyon ile sınırlı olmadığı ve glioblastomların %90 ında bulunduğu gösterilmiştir. Bu veri, glioblastomlarda gelişen Akt aktivitesinde PIKE-A nın önemli bir rolünü işaret etmektedir. Buna karşın, PIKE-S ve PIKE-L glioblastomlarda, neoplastik olmayan beyin dokusuyla karşılaştırıldığında düşük seviyelerde ifade edilir. İlginç olarak, yeni bir çalışma NF2 tümör süpresör gen ürünü merlinin Pi3k yı PIKE-L ye bağlanarak inhibe ettiğini gösterdi (281). PIKE-L ekspresyonunun susturulmasının merlinin tümör süpresör özelliğini ortadan kaldırdığını gösterdiler (281). Böylece azalan PIKE- L ekspresyonunun merlinin büyüme inhibitör etkisini azaltarak glioblastom patogenezine katkıda bulunabileceği düşünülebilir MDM2 [MDM2 p53 Binding Protein Homolog (Mouse)] MDM2 geni 12q13-14 kromozom bölgesinde yerleşmiştir (240). Buradan kodlanan MDM2 proteini (pmdm2), p53 ü kontrol altında tutar ve p53 ün G1/S geçişinde siklusu durdurma ve apoptoz etkisini engeller (255). MDM2 proteini sitoplazmada p53 e bağlanarak ve E3 ubiquitin ligaz aktivitesi ile p53 ü yıkarak p53 e karşı çalışan bir proteindir. Hipoksi, ultraviyole, radyasyon gibi hücrenin strese maruz kaldığı durumlarda pmdm2 nin p53 e bağlanma yerinde asetilasyon ve fosforilasyon nedeniyle yapısal değişiklik oluşur. Bu yüzden MDM2 proteini p53 ü bağlayamaz ve serbest kalan p53 transkripsiyonel aktivitesini göstererek G 1 ve G 2 kontrol noktalarında siklusun durdurulması ve bax geni aktivasyonu ile apoptoza neden olur. Ancak özellikle onkojenik stimulus varlığında MDM2 etkisini, p53 e bağımlı olmayan yolla gösterir. G1/S geçişinden sorumlu olan E2F/DP1 aktivasyonunu p14arf in inaktivasyonunu yapar. İnaktive olan p14arf hücre 51
73 çekirdeğinde MDM2 yi bağlayamaz ve sitoplazmaya çıkmasına engel olamaz. Bu nedenle P53 ün etkisi azalır. Ayrıca E2F aktivasyonu yaptığından hücre G1 fazından S fazına geçer (62). Bunun yanı sıra, MDM 2 nin p53 ten bağımsız etkileri son yıllarda büyük ilgi çekmektedir. MDM 2, p19/14arf, E2F, ribozomal proteinleri ve çeşitli hücresel proteinlerle etkileşim içerisindedir. MDM 2 onkoproteini birçok insan malinitesinde aşırı eksprese olmuş durumdadır ve yüksek MDM 2 seviyesi zayıf klinik prognoz ile ilişkilidir. MDM 2 ekspresyonunun antisens inhibisyonu, meme, kolon ve prostat kanseri modellerinde dayanıklı anti-tümör aktiviteleri ile sonuçlanmıştır (148). MDM2 genindeki amplifikasyon glioblastomlarda %10 un altında bulunmuştur (274, 275). İmmünohistokimyasal çalışmalarda MDM2 nin aşırı ekspresyonu primer glioblastomda %52, sekonder glioblastomda %11 bulunmuştur (23). MDM2 nin aşırı ekspresyonu, MDM2 geninin amplifikasyonu ve p53 mutasyonu bulunmayan primer glioblastomlarda saptanmıştır (274). Ayrıca kromozom 12q22 23 bölgesindeki heterozigosite kaybı glioblastomlarda %42 düzeyine kadar ulaşmaktadır. Bu bölge apoptoz aktive edici faktör 1 (Apaf-1) i içermektedir. Bu bölgede olan kayıplar nedeniyle Apaf-1 in fonksiyon görmemesi sonucunda p53 ün neden olduğu apoptoz yolunun sonlanması ile glioma onkogenezi başlamaktadır. Kromozom 12q bölgesindeki heterozigosite kaybının, p53 gen mutasyonu ve EGFR gen amplifikasyunu ile beraber olmaması, astrositik öncü hücrelerinin de novo olarak direkt malin transformasyonla primer glioblastom gelişiminden sorumlu tutulmaktadır (352). 52
74 OLIG2 (Oligodendrosit Transkripsiyon Faktörü 2) OLIG1 ve OLIG2 oligodendrosit hücrelerinde eksprese edilen temel heliksilmik-heliks olarak tanımlanmışlardır ve OLIG2 oligodendrosit gelişiminde erken markır olarak düşünülmektedir (194, 380). Fakat, OLIG2 ekspresyonu sadece oligodendroglial tümörlerde değil aynı zamanda astrositik tümörlerde de saptanmıştır (33, 193, 221, 239, 370). Diferansiyasyon inhibitörü proteinleri (ID); ID1, ID2, ID3, ve ID4, temel DNA bağlanma dizilerinin esikliği dışında, bhlh faktörlerine yapısal olarak benzeyen faktörlerdir (286). ID ler dominant negatif regülatörler gibi OLIG2 yi içeren bhlh faktörlerinin fonksiyonlarını onların heterodimerizasyonlarını bloke ederek ve sonra DNA dizilerine bağlanarak inhibe ederler (289). Dokuda yayılmış olarak bilinen astrositomlar, oligodendrogliomlar ve oligoastrositomlar en sık görülen primer beyin tümörleridir. Bu tümörler, farklılaşmış astrositlere ve oligodendrositlere histolojik benzerlikleriyle sınıflandırılırlar; ancak, bu yaklaşım, modern tedavi ve araştırmaların yönlendirilmesinde büyük kısıtlamalarla karşılaşmaktadır. Linaj markırları, morfolojik sınıflandırmaya potansiyel olarak faydalı bir ek sunmaktadır. Murin bhlh trankripsiyon faktörleri olan Olig1 ve Olig2 nöral progenitörlerde ve oligodendroglia da eksprese olmaktadır ve oligodendrosit gelişimi için gereklidir. Tek başına yüksek OLIG ekspresyonunun, oligodendrogliomları, astrositomalardan ayırdığı düşünülmektedir. OLIG2, bir sınıf olarak yayılmış tümörler için kullanışlı bir markırdır. Ancak, astrositomlarda OLIG2 nin ekspresyon heterojenliği, sadece OLIG2 ile yapılan bireysel gliomların immunohistokimyasal sınıflndırılmasını engellemektedir (185). 53
75 GSTT1 (Glutatyon S-Transferaz Teta 1) Glutatatyon S-transferazlar (GSTler), elektrofilik bileşikleri glutatyona konjuge etme yeteneğindedirler ve birçok metabolik süreçte temel rol oynamaktadırlar. Glutatyon S-transferaz T1 (GSTT1) sitozolik GST lerin teta sınıfının bir üyesidir (110, 111). İnsan ve hayvan dokularında karakterize olmuştur (201, 202, 215, 253, 262, 299, 305, 306). İnsan (h) GSTT1 geninde delesyon polimorfizmi bilinmektedir ve tüm beyaz ırkın yaklaşık olarak %20 si bu delesyonun homozigot taşıyıcısıdırlar. Bu bireyler GSTT1 fonksiyonundan yoksunlardır ve glutatyonu GSTT1 substratlarına bağlayamazlar. Bu nedenle; konjugatör olmayan olarak adlandırılırlar (253, 112). Endüstride kullanılan halojenlenmiş organik bileşikler ya da etilen oksit gibi ürünlere ek olarak, GSTT1 de BCNU yu [1, 3-bis (2-kloroetil)-1-nitrozüre] etkin bir şekilde metabolize edebilir. BCNU, nitrozüre grubunun sitostatik bir ilacıdır. Bunlar, malin gliomların kemoterapisinde kullanılan ilaçlardır. Glial tümörler arasında, glioblastoma multiformları, gliomların %50 sini oluşturmaktadır. Glioblastomalar, oldukça yüksek lokal invaziv olma özelliği gösteren astrosit tümörlerdir. Glioblastoma dokularının cerrahi nakli, genellikle intrakranial basıncın azalmasını sağlar. Bu tümörün yüksek invaziv olma özelliğinden dolayı, antimigratör ajanların konvensiyonel radyo-ve kemoterapiye eklenme olasılığı tartışma konusudur. Ayrıca, doğrudan neoplastik hücreleri hedeflemek için, intrakraniyal implantların kullanımının, glioblastoma hastalarında operasyon sonrası yaşamı olasılığını arttırdığı kanıtlanmıştır. Glioblastomaların sistemik kemoterapisi için, nitrozürelere ek olarak, örneğin; temozolimid gibi diğer bir alkilleyici ajanın kullanımı giderek artmaktadır. Ancak, glioblastomların kemoterapisindeki başlıca problemlerden bir 54
76 tanesi kemorezistanstır. Moleküler düzeyde, astrositlerin alkilleyici ajanlara olan duyarlılığının, DNA-tamir enzimi olan O6-metilguanin-DNA-metiltransferaz ın (MGMT) ve glutatyon sentezinin bloklanması ile büyük ölçüde arttığı in vivo olarak gösterilmiştir. MGMT nin, glioblastoma hastalarında, kemoresistansın temozolimid indüksiyonundaki rolü; Hegi ve arkadaşlarının yaptığı çalışmayla vurgulanmıştır. Glutatyon bağımlı yolak, mu ve pi alfa sınıfından olan glutatyon transferazların rolü ve ekspresyonu, glial tümörlerde büyük oranda incelenmiştir. Buna karşın, malin tümörlerde, teta sınıfı glutatyon transferazların ekspresyonu hakkında hiçbir bilgi yoktur. GSTT1 in BCNU yu büyük bir verimlilikle metabolize ettiği ve glioblastomalardaki modern teröpatik yaklaşımlardan birinin de BCNU nun lokal kullanımı olduğu gerçeği, GSTT1 in konjugatörlerde yapılan kemoterapide negatif bir etkiye sahip olduğunu düşündürmektedir. Eğer, GSTT1 tümör dokularında aktif halde olsaydı, bu durum mümkün olabilirdi. Ancak; bu soru hala cevaplanmamıştır. Günümüzde yapılan çalışmalarda, kemorezistansta GSTT1 in biyolojik anlamlılığını açığa çıkarmak amacıyla ilk olarak, insan malin gliomların neoplastik hücrelerinin GSTT1 i eksprese edip etmediği sorusuna yanıt bulmayı hedeflemişlerdir. Bu amaçla, bir astrosit ve oligodendrositik tümör tipinde HGSTT1 in ekspresyonunu araştırmışlardır (66) COX 2 (Siklooksigenaz 2) Prostaglandin endoproksit sentaz genel olarak siklooksigenaz (COX) olarak tanımlanır ve araşidonik asidin prostaglandinlere dönüşümü için gerekli olan anahtar bir enzimdir. Özellikle siklooksigenazın iki izoformunun (COX-1 ve COX-2) prostaglandin metabolitlerinin sentezini katalizlediği tanımlanmıştır (316). Fakat, 55
77 aralarında bazı farklılıklar fark edilmiştir (245). Gerçekte; genler farklı kromozomlarda lokalizedirler (COX-1 9q da ve COX-2 1p de), COX-2 nin aktif bölgesi COX-1 den daha büyüktür ve COX-1 in tersine COX-2 glukokortikoidler tarafından inhibe edilir. COX-1 (66 kd protein) neredeyse tüm normal dokularda eksprese edilir ve fiziksel homeostazda destek olur. Bunun tersine COX-2 (70 kd protein) sitokinler ve endotelin-1 tarafından indüklenebilir ve çeşitli uyarıcılara karşı cevabı artırır. COX-2 nin indüksiyonu, iskemi, travma, ensefalitis ve Alzheimer gibi birçok beyin hastalıklarında belgelenmiştir (17, 81, 98, 128, 196). Ayrıca, birçok çalışma COX-2 overekspresyonunu pankreatik, gastrik, prostatik, akciğer, baş ve boyun gibi birçok insan tümöründe (222, 350, 372) ve glioblastom gibi intrakranial neoplazmlarda (140, 309), ve meningiomlarda (186) tanımlamıştır. Bunun yerine COX-2 nin rolü kolorektal kanserde tanımlanmıştır (72). Nonsteroidal antiinflamatuar ilaçların potansiyel etkileri bulunmaktadır (138). Gerçekte, COX-2 anjiogenezin uyarılması, neoplastik hücrelerin invaziv potansiyelini regüle etme ve apoptoz gibi birçok tümör ilişkili prosesin içinde görülür. Tabiki; COX-2 aracılı yolaklarda diğer moleküller pozitif veya negatif sonuçlarla etkileşebilir. Örneğin artan bir COX-2 seviyesi bir antiapoptotik faktör olan bcl-2 nin artan ekspresyonu ile ilişkilendirilmiştir (337).Diğer taraftan tümör süpresör geni p53 ün COX-2 ekspresyonunu inhibe ettiği görülmüştür (324) htert (İnsan Telomeraz Revers Transkriptazı) Bir ribonükleoprotein enzim olan telomeraz, tümörlerdeki ayrıcalıklı ekspresyonundan dolayı, kanser tedavisi için potansiyel bir hedef olarak 56
78 düşünülmektedir. Özellikle malin gliomlar, telomeraz-hedefli tedavi için en iyi adaylardan bir tanesidir. Bunun nedeni, normal beyin dokuları telomerazı ekprese etmezlerken, malin gliomların baskın olarak telomeraz-pozitif olmalarıdır. Teorik olarak, telomeraz-pozitif tümörler için, telomeraz-ilişkili iki teröpatik yaklaşım bulunmaktadır. Bu yaklaşımlardan bir tanesi; doğrudan telomeraz aktivitesinin inhibisyonunu sağlayan ve apopotik hücre ölümü ya da büyüme aresti ile sonuçlanan anti-telomeraz kanser tedavisidir. Telomeraz holoenzim kompleksinin iki ana alt birimi olan RNA kalıbı (hter) ve katalitik alt birim (ters transkriptaz, htert) çok iyi birer teröpatik hedef olarak düşünülmektedirler. Diğer yaklaşım ise; telomeraz eksprese eden tümör hücrelerini hedefleyerek hücreleri doğrudan öldüren telomeraz spesifik kanser tedavisidir. Genellikle, telomeraz-pozitif hücrelere yönlendirilen immuntedavi gibi htert promotör sisteminin yönetiminde teröpatik genlerin transferini kullanan stratejiler bulunmaktadır. Bu telomeraz-ilişkili tedaviler, malin gliomların tedavisi için çok büyük umut vaat etmektedirler (160). Telomerler, lineer kromozomun uçlarında bulunan DNA sekanslarıdır (26). İnsanlarda 46 kromozom bulunur ve bu da, binlerce TTAGGG nükleotit tekrarını içeren 92 telomer ucu olduğu anlamına gelmektedir (26). Telomerlerin esas görevi, kromozomların uç bölgelerini herhangi bir hasara karşı korumak ve her DNA replikasyonundan sonra vital genetik bilgi kaybını engellemektir. Telomer bütünlüğü kromozom stabilitiesi için oldukça önemlidir ve telomerler tekrarlayan yaşlanma ve kanser oluşumunda büyük rol oynamaktadırlar (70). Telomerik sekanslar, DNA nın her replike oluşunda kısalırlar ve belirli bir boya indiklerinde hücre bölünmesi durur (241). Bu hücreler vücutta yıllarca yaşayabilir ancak, kendilerini replike edemezler. 57
79 Bu durumda, eğer telomeraz aktive olursa, bu aktivasyon telomerik sonun uzamasıyla sonuçlanır (27). Normal hücreler progresif olarak, kültürdeki pasajlar ile telomerik DNA larını kaybederken, neoplastik hücrelerde telomer uzunluğu stabildir (236). İnsan telomeraz RNA (htr), insan telomeraz ilişkili protein (htep1) ve insan telomeraz revers transkriptazı (htert) insan telomeraz kompleksinin bileşenleri olarak tanımlanmaktadırlar (236). Bu bileşenler arasında htert, anahtar role sahiptir çünkü ektopik ekspresyonu telomeraz aktivitesini yeniden düzenleyebilmek için yeterlidir (56). Bu nedenle, eğer bir hücrede telomeraz aktivitesi pozitif ise, hücrenin yüksek oranda proliferasyon göstermesi beklenir. Telomeraz birçok kanser tipinde aktif durumda olduğu için, bu kanser hücrelerinde malin transformasyon işaretlerinin görünmesi beklenir (149). Solid ve hematopoetik kanserlerde yapılan araştırmalarda, telomeraz ekspresyonunun kanser tedavisinde yeni bir hedef olduğu ve çok önemli bir diagnostik parametre olacağı yolundadır (208, 217). Servikal, kolon, over ve renal karsinomlar gibi birçok epitelyal kanser türü üzerine yapılan çalışmalarda, htert mrna ekspresyonu ve telomeraz aktivitesi arasında güçlü bir korelasyon gözlemlenmiştir. Bunun yanı sıra, normal olarak telomeraz negatif insan hücrelerinde htert geninin ektopik ekspresyonu, enzimatik aktivitenin yeniden oluşturulması ve hücresel yaşam süresini uzatmak için yeterlidir. Buna karşın, meme kanseri için telomeraz aktivitesi çelişkili sonuçlar vermiştir. Bununla birlikte, indirekt veriler telomeraz ve meme kanseri arasında bir korelasyon ileri sürmektedirler. Meme karsinomlarında sıklıkla bulunan farklı markır ekspresyonlarının telomeraz aktivitesinin kontrolünde önemli bir rol oynadığı 58
80 gösterilmiştir. Li ve arkadaşları, telomeraz aktivitesinin bir ihtimalle proliferasyon artışına neden olarak p53 ün down regülasyonu ile kontrol edilebileceğini göstermişlerdir. Bu durum başka bir çalışma ile de desteklenmiştir: yabanıl tip p53 ün htert mrna ekspresyonunu down regüle ettiği gösterilmiştir. Aynı şekilde, c- Myc onkogeni promotör bölgesindeki E-box a doğrudan bağlanarak htert transkripsiyonunu aktive etmektedir (20) Epigenetik Epigenetik terimi gen ekspresyonundaki DNA dizisinde tek başına kodlanmayan tüm mayotik ve mitotik kalıtım değişikliklerini tanımlar. DNA metilasyonunu, RNA ilişkili susturulmayı ve histon modifikasyonunu içeren 3 sistem epigenetik susturulmayı başlatmak ve güçlendirmekte kullanılır. Bu komponentler arasındaki ilişkiyi çözmek onların birbirini nasıl etkilediği ve stabilize ettiğini gösteren yeni kavramların oluşmasını sağlamıştır. Bu bir veya diğer birbirini etkileyen sistemlerdeki bozulma, epigenetik hastalıklarla sonuçlanabilecek, genlerin uygunsuz ekspresyonu veya susturulmasına yol açar. DNA daki sitozin rezidülerinin C 5 pozisyonunun metilasyonu epigenetik susturulma mekanizması olarak tanımlanmıştır. İnsan genomundaki CpG bölgelerinin metilasyonu bir seri DNA metiltransferaz (DNMTs) ile korunmaktadır ve aktarılabilir öğelerin susturulmasında, viral dizilere karşı savunmada ve belli genlerin bastırılmasında transkripsiyonel rolü vardır. 5-Metilsitozin, insan DNA sında CpG metil akseptör bölgesinin güçlü bir baskılanması ile sonuçlanan C:G T:A transizyonuna sebep olacak kadar yüksek 59
81 derecede mutajeniktir. Büyüklükleri 500 baz çiftinden daha fazla bölgeler olan ve %55 ten daha çok GC içeriğine sahip olan CpG adaları evrim boyunca korunmuştur, çünkü metilasyondan ayrı kalırlar. DNA nın bu bölgeleri memeli genlerinin %40 oranında promotör bölgeleri içinde yer alırlar ve metilendikleri zaman stabil olarak kalıtılan transkripsiyonel susturulmaya sebep olurlar. CpG adalarının sapmış de novo metilasyonu insan kanserlerinde bir özelliktir ve erken dönem karsinogenezinde yer alır. Histon modifikasyonları epigenetik modifiyeciler olarak tanımlanmıştır. Asetilasyonu, amino asit terminal uçlarındaki korunmuş lizin rezidülerinin metilasyonunu içeren histonların post translasyonal modifikasyonları birkaç yıldır çalışılmaktadır. Genellikle hipoasetile histonlar transkripsiyonel olarak inaktif ökromatik veya heterokromatik bölgelerde bulunurken, histon asetilasyonları aktiftir. Kromatinin hem aktif hem de inaktif bölgelerinde, histon metilasyonu bir markır olarak kullanılabilir. Histon H3(H3-K9) un N terminusu üzerindeki lizin 9 un metilasyonu sessiz DNA nın bir özelliğidir ve sentromerler ve telomerler gibi heterokromatik bölgeleri düzenler. Ayrıca inaktif X kromozomu ve susturulmuş promotörler üzerinde bulunur. Buna karşın Histon H3(H3-K4) un lizin 4 ünün metilasyonu aktivitenin işaretidir ve aktif genlerin promotör bölgelerinde predominant olarak bulunur. Çünkü lizin metilasyonu monomerik, dimerik veya trimerik olabilir ve histonlar fosforilasyon gibi diğer posttranslasyonal modifikasyonlara bağımlı olabilirler. Bu geniş varyasyon farklı modifikasyonların olası kombinasyonlarının çeşitliliğine sebep olabilir. Bu, farklı selüler faktörler tarafından okunabilen ve değerlendirilebilen bir histon kodunu oluşturabilir. Histon modifikasyonları ve DNA metilasyonu arasındaki bağlantılar bitkilerde ve mantarlarda H3-K9 metilasyonunun DNA metilasyonu için gerekli olduğu bölgelerde 60
82 bulunmuştur. DNA metilasyonu ayrıca H3-K9 metilasyonunu başlatır ve ayrıca memelilerde H3-K9 metilasyonununun DNA metilasyonunu perisentromerikten heterokromatine yönlendirdiği bölgelerde gösterilmiştir. Histon deasetilaz (HDACs), histon metiltransferaz ve metilsitozin bağlayıcı proteinler arasındaki ilişki DNA metiltransferaza sebep olur. Post transkripsiyonal susturulmada RNA nın rolü ilgi uyandırmıştır. Buna karşın antisens transkriptlerindeki RNA, kodlanmayan RNA lar, RNA interferans heterokromatin formasyonu ile mitotik olarak kalıtılan transkripsiyonel susturulmaya sebep olabilir. RNAi, DNA metilasyonu ve H3-K9 metilasyonu arasındaki benzer bağlantı Arabidopsis thaliana da gösterilmiştir. RNAi yönlendirilmiş heterodimerik bölgelerin susturulması memelilerde henüz gösterilmemiş olmasına rağmen RNA nın farklı susturma mekanizmaları tanımlanmıştır. Örneğin; antisens RNA lar bazı memeli imprintlenmiş genlerinin susturulmasında mevcuttur. Talasemiye ait yeni bir durum raporunda antisens transkripsiyonunun DNA metilasyonuna ve globin geninin stabil susturulmasına neden olabileceği gösterilmiştir. Bu yüzden, RNA, histon mdifikasyonuna (H3-K9 metilasyonu) yönelmede ve spesifik lokuslar için (perisentromerik heterokromatin) DNA metilasyonuna anahtar bir yol açıcı olabilir, bu suretle kalıtımsallık ve stabil susturulmayı çağrıştırabilir. Kansere yol açan yolakların bozulmaları genelde ilgili genin promotörünün de novo metilasyonu sebebiyledir. Epigenetik susturulma Knudson hipotezini karşılayan üçüncü yolak olarak kabul edilmiştir. Kromatin modifiye enzimler farklı hemapatolojilerin etiyolojisi ile ilişkilendirilmiştir. İnsan lösemisinin bir karakteristiği çeşitli kromozomal translokasyonların varlığının, füzyon proteinleri ekspresyonuna 61
83 yol göstermesidir. Hem histon asetiltransferaz hem de histon metiltransferaz füzyonların bir parçası olabilir ve hedef genlerin upregülasyonuna sebep olabilir. Akut promiyelositik lösemide onkogenik füzyon proteini PML-RAR (promiyelositik lösemi-retinoik asit reseptörü) hemopoietik hücrelerin diferansiyasyonu için gerekli genleri baskılamak için bir HDAC çalıştırır. Benzer şekilde, akut myeloid lösemide AML1-ETO füzyonları baskılayıcı N-CoR Sin3 HDAC1 kompleksini çalıştırır ve myeloid gelişimini inhibe eder. Son zamanlarda hastalıkta ATP bağımlı kromatin biçimlenmesinin rolü ile doğru kromatin kompozisyonunun altı çizilmiştir. Bunlar nukleozomları hareketlendirmede ve kaydırmada ve böylelikle transkripsiyonu regüle etmede yetenekli çoklu alt ünitelerdir. Yüksek korunmalı SWI-SNF komplekslerinin birçok üyesi kanserde görülür. Örneğin, SNF5 kaybı pediyatrik kanserlerde gözlenmiştir ve ATPaz altüniteleri BRM ve BRG1 çeşitli kanser hücre hatlarında ve primer tümörlerde mutasyona uğramıştır, bu da küçük olmayan akciğer kanserli hastaların zayıf prognozu ile ilişkilidir Epigenetik Yığılma Epigenetik değişiklikler ile genetik değişiklikleri karşılaştırmanın ayırt edici bir özelliği, beklenmeyen bir işlemden ziyade aşamalı bir kazanılmış işleme eğilimli olmalarıdır. Örneğin, genomik 5-metilsitozin konsantrasyonlarındaki bir düşüş, memeli hücre yaşlarında meydana gelir ve DNA metilasyonunun bu aşamalı kaybı anormal gen aktivasyonu ile sonuçlanabilir. 5-metilsitozindeki düşüş gen promotörlerindeki CpG adalarının lokalize hipermetilasyonu ile aynı zamanda meydana gelir. Metile CpG lerin promotör içinde birikmesi, gen susturulmasında şalter yerine daha çok bir direnç aygıtı gibi rol oynar. Bu değişimler önleme 62
84 stratejileri için hedeftirler. DNA metilasyonunda veya DNA metil bağlı proteinlerdeki bir düşüş, kansere eğilimli farelerde intestinal poliplerin sayısını azaltabileceği gösterilmiştir. DNA metilasyonu, prokaryot ve ökaryotlarda meydana gelen bir olaydır. Prokaryotlardaki DNA metilasyonu, kendi sınırlayıcı enzimleri tarafından yabancı DNA yı elimine ederek sahip olduğu DNA yı korur. Yüksek ökaryotlarda DNA metilasyonu, gen eksprasyonunu düzenlemek için diğer bir metot gibi davranır. Yüksek ökaryotlarda DNA metilasyonu, sapmış metilasyonun, kanserde yaygın bir olgu olduğu ve en erken değişikliklerin arasında onkogenez esnasında meydana gelebildiği gösterildi. DNA metilasyonu, gen imprinting, embriyonik gelişim, X kromozom gen susturulması ve hücre döngüsünü düzenlenmesinde merkezi rol oynayıcı olarak gösterilmiştir. Bazı bitki ve hayvanlarda, DNA metilasyonu, bir metiltransferaz enzimi yoluyla sitozin pirimidin halkasının beşinci karbon pozisyonuna bir metil grubunun eklenmesine dayanır. DNA metilasyonun çoğunluğu memelilerde 5 - CPG 3 dinukleotidleri içerir fakat diğer metilasyon kalıpları vardır. Gerçekte bütün 5 - CPG 3 dinukleotidlerinin %80 civarı memeli genomlarında metilenmiş olduğu bulunmuş, öyleki metile olmayanların %20 si promotör bölgelerde ya da genin ilk ekzonunda bulunur. DNA metilasyonunu nicelemek ve belirlemek, kanser, gen ekspresyonu, genetik hastalıkların çalışılmasının ve ayrıca daha birçok önemli biyolojik çalışmanın olmazsa olmazı haline gelmiştir. Bu güne kadar DNA metilasyonunu ölçmek için yüksek-performanslı kapiller elektroforezi ve metilasyon hassaslı PCR gibi birçok metot geliştirilmiştir. Şu an ki en kullanılır yöntem bisülfit metodudur. Bu teknik, 63
85 metile sitozinler değişmeden kaldığında urasile dönüşecek metile olmayan sitozinlere neden olan DNA nın bisülfit ile muamele edilmesidir. Bu protokolde bisülfitlenmiş DNA, PCR ile çoğaltılır ve PCR ürünleri DNA dizi analizi ile analiz edilir ya da restriksiyon endonükleaz ile kesilir. Daha sonra DNA segmentinin metillenmiş profili muamele edilmemiş DNA ile bisülfitlenmiş DNA sekanslarının karşılaştırılması ile belirlenmektedir (73). Hipermetile promotör bölgelerinin tümör belirleyicisi olarak kullanımı rutin olarak kullanılmasa da pekçok yönden avantajlıdır. Kanser türlerinin çoğunda, CpG adacık hipermetilasyonu tümör tipine özgü izlenmektedir. Ayrıca, promotör hipermetilasyonu kanserlerin erken döneminde oluştuğu için erken tanıya da olanak sağlamaktadır (83) Metilasyon Biyokimyasal olarak tümör baskılayıcı genlerin metilasyonu, delesyonlara karşı alternatif bir mekanizma oluşturmaktadır (7). Günümüzde tümör baskılayıcı genlerin heterozigosite kayıplarında diğer alelin en azından metilasyona uğradığı düşünülmektedir. Metilasyon; CpG dinükleotitinde, sitozin rezidülerinin, metil grubu ile birleşmesi ile gerçekleşir ve tümör baskılayıcı genlerin ve hatta bir kromozomun fonksiyon kaybı ile sonuçlanır (X inaktivasyonu gibi) (292). Astrositom onkogenezinde on altı genin promotör-cpg alanlarının metilasyonu bulunmuştur (374). Glioblastomlarda PTEN promotör bölgelerinin metilasyonu %35 olarak rapor edilmiştir (16). p16 promotör geninin hipermetilasyonu, bu genin inaktivasyonuna neden olmaktadır ve tüm gliomlarda % 24 düzeyinde bulunmuştur (54, 89). 64
86 Nitrozüre ve temozolomid, antikanser etkisini DNA daki O6- guanini alkilleyerek yapmaktadır. O6- metilguanin DNA metiltransferaz (MGMT) ise O6- guanini tekrar tamir etmektedir. Bu nedenle O6-metilguanin DNA metiltransferaz (MGMT) alkilleyici ilaçlara ve temozolamide karşı direnç gelişiminden sorumludur (88). MGMT geninin promotör bölgesinin metilasyonu, tüm glioblastomlarda %45 düzeyinde bulunmuş ve bu bölgede metilasyon gösteren hastalar temozolomid tedavisine iyi yanıt vermiştir (29, 113). Öte yandan MGMT geninin promotör bölgesinin metilasyon yolu ile susması, primer glioblastomlarda %36 düzeyinde bulunmuştur (231). Son zamanlarda, kromozom 3p21.3 bölgesindeki RASSF1A geninin hipermetilasyonu beyin tümörleri dahil kanser vakalarında rapor edilmiş ve bu genin tümör baskılayıcı bir gene aday olduğu düşünülmüştür (122). Düşük evreli glial tümörlerde RASSF1A hipermetilasyonu % 31,7 bildirilmiştir (95). Bir çalışmada glioblastomlarda bu genin hipermetilasyonu 6/10 hastada saptanmıştır (71) Metilasyona Özgü PCR DNA metilasyonun saptanması DNA dizisindeki sitozin ile 5-metilsitozinin ayırt edilmesine dayanır. Metilasyonun saptanması için 3 temel strateji kullanılmaktadır: DNA nın metilasyona duyarlı bir restriksiyon endonükleaz ile kesilmesi (2); DNA nın sodyum bisülfat ya da metabisülfat ile kimyasal olarak modifikasyonu (279); 5-metilsitozinin imünopresipitasyonu ile genomun metilenmiş ve metillenmemiş bölgelerinin birbirinden ayrılması (73). Restriksiyon enzimlerinin kullanımına dayalı tekniklerden biri restriksiyon işaretli genom görüntüleme (RLGS) tekniğidir. 2 boyutlu bir yaklaşımla uygulanan RLGS, restriksiyon enzim 65
87 polimorfizmleri ve DNA metilasyon duyarlı bölgelerini kullanır. Ancak duyarlılığı düşük ve oldukça zaman alan bir yöntem olduğundan günümüzde pek tercih edilmemektedir. Daha başarılı bir yöntem olarak, metilasyona duyarlı NotI enzim kesiminden elde edilen fragmanların biyotinle işaretlenmesinden sonra BAC arraylerinde hibridizasyonu kullanılmıştır. Bu sayede farklı tip hücrelerle (astrosit ve periferal kan lenfositleri) yapılan çalışmalar sonucu insan, rat ve farelerde korunmuş olan bir metilasyon farklılığı saptanmıştır. Restriksiyon enzim kesimi ile microarrayleri birlikte kullanan diğer bir yöntem ise HELP ( HpaII ufak fragmanın zenginleştirilmesinin ligasyon aracılı PCR) olarak bilinir. Bu yöntemde, metile olmayan DNA ve genomik DNA nın farklı hibridize olmasından yararlanılır ve HpaII kesim fragmanlarıyla, metilasyona duyarlı olmayan MspI fragmanlarının birlikte hibridizasyonu ile gerçekleştirilir. DNA nın bisülfitle muamele edilmesi, sitozinin urasile dönüşmesine neden olur. Bu reaksiyondan yararlanılarak geliştirilen yöntemde, PCR dan sonra yapılan DNA sekans analizinde metillenmemiş sitozinler timin olarak okunur, metillenmiş olanlar ise sitozin olarak kalırlar, böylece metillenmiş ve metillenmemiş DNA saptanabilir. Üçüncü epigenomik yaklaşım ise ChIP (kromatin immunopresipitasyon) yönteminden yola çıkılarak geliştirilmiştir. Anti-metilsitozin antikorları kullanılarak metilenmiş DNA nın genomdan izole edilmesi sağlanır, ve MeDIP (metillenmiş DNA immunopresipitasyonu) olarak adlandırılır. ChIP, hücrenin kromatin durumunun saptanmasında yaygın olarak kullanılmaktadır. DNA ve ilişkili olduğu proteinlerin 66
88 kimyasal muamele ile çapraz bağlanmaları sağlanarak, DNA-protein kompleksleri aranan histon modifikasyonuna uygun bir antikor ile immunopresipite edilir. Bu yöntem mikroarray çipleriyle uygulandığında hızlı ve sağlıklı analizler yapılabilmektedir ve ChIP üzerinde chip olarak adlandırılmaktadır. Yöntem, uygulayıcısına esneklik sağlamakta ve spesifik ya da daha genel antikorlar kullanılarak istenilen bölgelerdeki histon modifikasyonları incelenebilmektedir Beyin Tümörü Modelleri Hücre hatları, laboratuar çalışmalarında ve in vitro kanser araştırmalarında model olarak sıklıkla kullanılmaktadır. Hücre hattı kullanımının bir takım avantajları bulunmaktadır. Muamele edilmeleri kolaydır ve sınırlı koşullarda bile limitsiz üreyebilme yeteneğindedirler. Ayrıca, yüksek derecede homojenite gösterirler ve donmuş stoklardan kolaylıkla kullanılır hale getirilebilirler. Ancak, bir takım dezavantajları da bulunmaktadır. Hücre hatları sürekli kültürde genotipik ve fenotipik sapmalara eğilimlidirler. Ayrıca, alt kültürler oluşabilir ve zaman içerisinde popülasyonda bulunan daha hızlı büyüyen ve spesifik klonların seçimiyle fenotipik değişikliklere neden olabilirler. U-87 MG Hücre Hattı Organizma: Homo sapiens (insan) Yaş: 44 Cinsiyet: Bayan Etnik köken: Beyaz Irk Organ: Beyin Büyüme Özelliği: Adheren Morfoloji: Epiteliyal Hastalık: Glioblastom, astrositom Tümör derecesi: 2007 de derece IV olarak sınıflandırıldı Morfoloji: Epitelyal Tumorigenik: Evet 67
89 Antijen ekspresyonu Kan tipi A, Rh+ Hücrelerin % 48 inde rastlanan 44 kromozoma sahip hipodiploid insan hücre hattıdır. Yüksek kromozom düzenlenme derecesi % 5,9 dur. 12 markır der(1)t(1;3) (p22;q21), der(16)t(1;16) (p22;p12), del(9) (p13) ve başka Sitogenetik Analiz: 9 tane dahil olmak üzere tüm hücreler için ortaktır. Birçok hücrede der(1) markırı 2 kopyaya sahiptir. Normal X ten sadece 1 kopya vardır. N1,N6 ve N9 bulunamadı. İzoenzimleri: AK-1, 1 ES-D, 1 G6PD, B GLO-I, 1 Me-2, 1 PGM1, 2 PGM3, 1 U-118 MG Hücre Hattı: Organizma: Homo sapiens (insan) Yaş: 50 Cinsiyet: Erkek Etnik köken: Beyaz Irk Organ: Beyin Büyüme Özelliği: Adheren Morfoloji: Karışık Hastalık: Glioblastom, astrositom Tümör derecesi: 2007 de derece IV olarak sınıflandırıldı Tumorigenik: Evet Antijen ekspresyonu Kan tipi A, Rh+; HLA Aw24, A28, B12, Bw47 Amelogenin: X,Y CSF1PO: 11,12 D13S317: 9 D16S539: 12,13 DNA Profili D5S818: 11 (STR): D7S820: 9 THO1: 6 TPOX: 8 vwa: 18 AK-1, 1-2 ES-D, 1 İzoenzimleri: G6PD, B GLO-I,
90 Me-2, 1 PGM1, 2 PGM3, 2 LN-18 Hücre Hattı: Organizma: Homo sapiens (insan) Doku: serebrum; sağ temporal lob İzolasyon Tarihi: 1976 Tümör derecesi: Derece IV Hastalık: Glioblastoma; glioma Hücresel Ürünler: Fibronektin Yaş: 65 Cinsiyet: Erkek Etnik köken: Beyaz Organ: Beyin Büyüme Özelliği: Adheren Morfoloji: Epitelyal Tumorigenik: Evet Antijen ekspresyonu HLA A2, A9, B5, BW35, DRW3 Amelogenin: X,Y CSF1PO: 12 D13S317: 12,13 D16S539: 11,13 DNA Profili D5S818: 11,13 (STR): D7S820: 8,10 THO1: 9 TPOX: 8 vwa: 17,18 Onkogen: p53 + (mutasyona uğramış, TGT (Cys)--> TCT (Ser) kodon 238 de mutasyon), PTEN + (yabanıl tip), p16 -(silinmiş), p14arf -(silinmiş) DAOY Hücre Hattı: Organizma: Homo sapiens (insan) Organ: Beyin Doku: Serebellum İzolasyon Tarihi: 1985 Hastalık: desmoplastik serebellar medulloblastoma Hücresel Ürünler: fibronektin 69
91 Yaş: 4 Cinsiyet: Erkek Etnik köken: Beyaz Irk Büyüme Özelliği: Adheren Morfoloji: Poligonal Tumorigenik: Evet Uygulamalar: transfeksiyon konağı Amelogenin: X CSF1PO: 11 D13S317: 13,14 D16S539: 10 DNA Profili (STR): D5S818: 11,13 D7S820: 8,10 THO1: 9 TPOX: 8,10 vwa: 14,20 Hipertetraploid bir insan hücre hattıdır. Yüksek kromozom dizilim frekansına sahip hücreler %2 dir. Kök hücre hattı kromozom sayısı yüksek olduğundan beri poliploidi için değerlendirme değişebilir. Tüm hücrelerde 13 veya daha fazla marker kromozomu ortaktır. Bazıları hücre başına 2 den 4 e kadar kopya içerir. t(1q5q), t(13q;?), 15p+, 7q+, Sitogenetik Analiz: der(9)t(3;9)(p21;q34) and 8 tane daha markerlar arasındadır. Birçok hücrede 15p+ 3 kopya ve der(9) 4 kopya içermektedir. Bazı hücreler del(1)(p11) e sahiptirler. Normal N12,N14,N15 ve N19 hücre başına 4 veya daha fazla kopya içerir. çoğu hücrede 2 normal X kromozomu yer alır, fakat keşfedilebilir normal Y yoktur. İzoenzimler: İkilenme Zamanı: Comments: AK-1, 1 ES-D, 1-2 G6PD, B GLO-I, 1 Me-2, 1 PGM1, 2 PGM3, saat DAOY hücre hattı ilk kez Batı Avustralya daki Royal Perth Hastanesindeki P. F Jacobsen tarafından 1985 yılında keşfedildi. Hücre hattı 4 yaşındaki erkek çocuktan ön lobdan biyopsi ile alınan tümör hücre materyalinden elde edildi. Orjinal tümör nöronal ve glial farklılaşma karakterine sahipken bu özellikler hücre hattı tarafından gerçekleştirilmez. Hücrelerin dibutiril siklik AMP (camp) ile tedavisi bu karakterlerin ekspresyonunu içermediği S100 proteinlerinin ve glial fibriler asidik proteinlerin boyanması ile ölçüldü. 70
92 WI-38 Kontrol Hücre Hattı Organizma: Yaş: Cinsiyet: Etnik Köken Organ: Hücre Tipi: Hastalık: Virus Hassasiyeti: Revers Transkripti: DNA Profili (STR): Sitogenetik Analizi: İzoenzimleri: Homo Sapiens (İnsan) 3 Aylık Fetüs Bayan Beyaz Irk Akciğer Fibroblast Normal Veziküler Stomatitis; Herpes Simplex; Pseudorabies Poliovirus 1 Negatif Amelogenin: X CSF1PO: 10,12 D13S317: 11 D16S539: 11,12 D5S818: 10 D7S820: 9,11 THO1: 8,9.3 TPOX: 8 Vwa: 19,20 Normal Diploid G6PD, B Hs 605.Sk Kontrol Hücre Hattı: Organizma: Homo sapiens (insan) Organ: Deri Hastalık: Normal Yaş: 61 Cinsiyet: Bayan Etnik köken: Beyaz Irk Büyüme Özelliği: Adheren Morfoloji: Fibroblast Amelogenin: X CSF1PO: 12,14 DNA Profili (STR): D13S317: 9,12 D16S539: 10,11 D5S818: 11,13 D7S820: 10,11 THO1: 6,9.3 71
93 TPOX: 8 vwa: 15, Gen Ekspresyonu ve Gen Kopya Sayısı Gen ifadesi ya da gen ekspresyonu, DNA dizisi olan genlerin, fonksiyonel protein yapılarına dönüşmesi süreci için kullanılan bir terimdir. Basitçe, bu durum genlerin açık (aktif) olup olmadıkları olarak da tanımlanabilir. DNA'nın farklı genlere karşılık gelen bölgelerindeki baz dizilişleri, o gen tarafından şifrelenen proteinin aminoasit dizisini belirler. DNA'daki baz dizilişlerinin proteinin amino asit dizisini nasıl şifrelediği genetik ve biyokimyasal yöntemlerin birlikte kullanılması ile ortaya konulmuştur. İnsanda bulunan yaklaşık 30 bin genin hepsi, aynı anda aktif olarak bulunmayabilir. Gen ifadesinin her adımı, transkripsiyondan post-translasyonel modifikasyona kadar adımlarda değişebilir. Gen düzenlenmesi, hücreye; hücre farklılaşması, morfogenezis ve canlıdaki çok yönlü adaptasyonun temellerini oluşturan, yapı ve fonksiyonların kontrolünü verir. Protein olmayan genler, (rrna genleri, trna genleri, vs.) protein olarak translasyon edilmezler. Bazı genlerin ifadesi, (mikrornalar ya da ubiquitin ligazlar gibi) transkripsiyondan sonra düzenlenir, böylece artan mrna konsantrasyonu herzaman gen ifadesine gerek duymaz. Bununla beraber, mrna seviyeleri Northern 72
94 blot yöntemiyle nicel olarak ölçüldüğünde, RNA'nın tamamen bu ifadeye bağlı olduğu görülmüştür. Gen kopya sayısı (ya da kopya sayısı varyantları (CNV), kopya sayısı polimorfizmleri (CNP) bir bireyin genomunda bulunan bir genin kopya sayılarında bulunan farklılıklardan oluşan genetik karakter özelliklerini ifade eder. Bazı genlerin kopya sayılarında görülen farklılıkların insanlarda genetik çeşitliliğe katkıda bulunduğu gerçeği İnsan Genom Projesi nin tamamlanmasıyla ortaya çıkarılmıştır. Bazı insanlarda yalnızca bir kromozomda bazı genlerin delesyonu görülürken bazı insanlarda aynı genlerin birçok kopyası görülmektedir. Yaklaşık insan geni içinde, akraba olmayan insanlar tipik olarak yaklaşık 12 farklı yerde değişik kopya sayısına sahiptir ve büyük çaplı değişikliklerin boyutları birkaç kilobaz ile 2 megabaz arasında değişir (301). Nükleotid dizisi değişiklikleri insan genetik varyasyon çalışmalarının önemli bir bölümünü oluşturmaktadır. Bundan dolayı; genomik alanda oldukça büyük ilgi oluşturduğu kabul edilmektedir. Çeşitliliğin birincil tipi, tek nükleotid polimorfizmin seviyesi olarak keşfedilmiştir. Kayıp ve kazançların seviyesindeki geniş çeşitlilik, kopya sayısı çeşitliliği, copy number variation (CNV), olarak da isimlendirilmektedir. CNV ların genom genelinde tespiti, yüksek çözünürlüklü ve high-throughput teknolojilerin yoksunluğundan dolayı zor olmaktadır. Bu varyasyonları tanımlamadaki temel basamağı, microarray tabanlı karşılaştırmalı genomik hibridizasyonun (array-cgh) gelişmesi oluşturmaktadır (206, 256, 319). 73
95 Son zamanlarda, farklı yaklaşımlar kullanarak insan genomunda CNV lerin varlığını ortaya koyan iki önemli çalışma rapor edilmiştir (129, 301). Her iki çalışmada, segment dublikasyonu ve genler ile örtüşen genomik dengesizliğin normal bireylerindeki varlığı ikna edici bir şekilde gösterilmiştir ve bu da, hastalık olasılığı ve fenotipik varyasyona katkıda bulunabilir. Bu başlangıç bulguları, CNV nin önemine kanıtı güçlendirecek çok sayıda çalışma tarafından takip edilmektedir (270). DNA kopya sayısı değişimlerinin tanımlanması tümör progresyonu ve mekanizmanın anlaşılması kanser çalışmalarında büyük önem taşımaktadır. Yapısal yeniden düzenlenmeler ya da kazanımlar onkogenlerin aktivasyonuna neden olabilir, oysa delesyonlar tümör süpresör genleri inakive edebilir. Tüm bu anormallikler sözü geçen fenotip değiştirici genlerin ekspresyonuna da etki edebilir. Bu genler gibi tumor başlangıcında çok kritik olmasa da, hastalığın dışa yansımasında ve klinik resminde değişikliklere yol açabilir. Kopya sayısı değişimiyle etkilenmiş genomik bölgelerin fonksiyonel değerlendirilmesi ve keşfi, tanısal uygulamalar ve kanser biyolojisini anlamak için temel oluştırmaktadır. Tipik array-cgh denemesi, test ve referans örneklerinden toplam genomik DNA, mikroarraye hibridize edilmiş ve farklı bir şekilde işaretlenmiştir. Mikroarreyde her bir spottaki referans ve test sinyali arasındaki yoğunluk oranı, teorik olarak, ilgili genomik sekansın relatif kopya sayısı ile orantılıdır. Son zamanlarda ticari olarak geliştirilen ve sipariş üzerine üretilen mikroarreyler çok yüksek çözünürlükte kopya sayısı analizlerini mümkün kılar. Bu gibi deneylerden oluşturulan büyük miktarlardaki verilerin sonuçları DNA kopya 74
96 sayısı profilini test etmek için otomatik prosedürlerin kullanımı array-cgh verilerinin analizinde temel basamağı oluşturmaktadır. Son zamanlarda bu konu ile ilgili bir metot önerilmiştir. Metotların çoğunluğu bilinmeyen kesme noktasındaki varyans değişiklikleri vasıtasıyla Gaussian dağılımları ile bir model varsaymaktadır. Bu gibi kesim noktaları bulgusunun görevi sıklıkla segmentasyonu işaret etmektedir (357). Bu metotlarla paylaşılan yaygın yaklaşım, muhtemel dağılım fonksiyonu olasılığını maksimize eden tutum içinde kesim noktalarını tanımlamaktır (12, 126, 345). Diğer genetik varyasyonlar gibi, bazı gen kopya sayısı varyantlarının hastalığa yatkınlık ya da direnç gösterme gibi özelliklerinin bulunduğu görülmüştür. Gen kopya sayıları kanser hücrelerinde yüksek olabilir. Örneğin, EGFR kopya sayısı küçük hücreli olmayan akciğer kanserinde normalden fazla olabilir (45). Ayrıca CCL3L1 geninin yüksek kopya sayısının insan HIV enfeksiyonuna yakalanmada daha az duyarlı olduğu (100), ve FCGR3B geninin düşük sayısının sistemik lupus eritematoz ve benzeri yangısal bağışıklık sistemi bozukluklarına duyarlılığı artırabildiği (266) gösterilmiştir. İnsanlarda geniş çaplı kopya sayısı varyantlarının birçok tek nükleotid polimorfizmi kadar DNA yı kapsadığını ve benzer şekilde fenotipik çeşitliliğinden ve hastalık duyarlılığından sorumlu olabileceği belirtilmiştir (271) Eksklüzyon Boyaları Canlı (bütün plazma membranı olan) ve ölü (hasarlı plazma membranı olan) hücreler diferansiyel boyama ile ayırt edilebilir. Hasar görmüş plazma membranı 75
97 permeabilitesi olan hücreler boyanırken, hasar görmemiş (canlı) hücreler, boyalar plazma membranını geçemedikleri için bu boyalarla (eksklüzyon boyaları) boyanmazlar. Eksklüzyon testleri için en sık kullanılan boya tripan mavisidir Tripan Mavisi Boyası Canlılık Testi Tripan Mavisi Boyası sadece ölü hücrelerin membranlarından geçebilir. Her bir muamele edilen grubun yaşama yüzdesi ölü hücrelerin (mavi) ve canlı hücrelerin (boya almamış) sayılmasıyla belirlenir. Analiz edilen parametre; hasarlı, sızıntı yapan plazma membranıdır. Bu testin avantajları arasında ışık mikroskobu ile canlı hücrelerin ölülerden ayırt edilmesinde kullanılan standart bir metot olması, hızlı ve ucuz olması, total hücre popülasyonundan çok küçük bir hacmin test analizi için gerekli olması sayılabilir. Sayım için en sık kullanılan yöntem Neubauer hemositometresidir ve iki eşdeğer bölmeye sahiptir (Şekil 3). Sayma odacığının derinliği 0.1mm dir. Hücre süspansiyonu, odacığa mikro pipetler yardımıyla aktarılır. Mevcut hücre sayısına göre hücreler, karelerin üçlü çizgilerle çevrelenmiş olduğu merkezi bölmelerde sayılır. Sol ve üst üçlü çizgilerdeki hücreler sayıma dahil edilir. Bu bölmelerin alanı 1 mm 2 dir. Böylece hacmi 0,1 mm 3 olur. Bu alandaki tüm hücreler sayılırsa (N): N=0.1mm 3 te mevcut hücre sayısı 1 ml yani 1cm 3 teki hücre sayısı = N х DF x 10 4 olur. DF=Dilusyon Faktörü 76
98 Şekil 3: Neubauer hemositometresi (382) 77
99 BÖLÜM II 2.1. GEREÇ VE YÖNTEM Ege Üniversitesi Nöroşirurji Anabilim Dalı nda yılları arasında beyin tümörü tanısı almış 98 olguda, 7 onkogen ve 7 tümör süpresör genin ekspresyonları, kopya sayısı aberasyonları ve tümör süpresör genlerin promotör bölgelerinin metilasyonları tanımlayıcı ve prospektif olarak çalışılmıştır Hücre Kültürü ve Moleküler Çalışmalarda Kullanılan Cihazlar ve Kimyasallar Tablo 1: Kullanılan cihazlar CİHAZLAR MARKA CO 2 li inkübatör Thermo Otomatik mikropipetler Eppendorf (10, 20, 100, 200, 1000 µl lik) Laminar air Flow Heraus İnverted Mikroskop Olympus, Japan Işık Mikroskobu Olympus, Japan Thermomixer comfort Eppendorf 1.5ml Steril kültür kapları (flask) 96 kuyucuklu hücre kültür kabı Corning Corning Neubauer Lam Marienfeld, Almanya Scientific Finnpipette Thermo PCR TC-512 Techne Hassas Terazi Sartorius GD 603 Nanodrop Spectrophotometer ND-1000 IEC micromax Microcentrifuge Thermo Real Time online PCR Roche Diagnostics, Germany
100 Tablo 2: Kullanılan kimyasal maddeler Madde RPMI 1640 Dimetil sülfoksit (DMSO) Tripan Mavisi Boyası (%0,5 (w/v) fizyolojik tuz içinde) Fötal Sığır Serumu (sıcaklık ile inaktive edilmiş) Penisilin/ Streptomisin Gentamisin BIOAMF Medyum DMEM Medyum EZ DNA Methylation Gold Kit Transcriptor First Strand cdna Synthesis Kit Kollegenaz Tip IV High Pure RNA Isolation Kit Taqman Probe Master High Pure PCR Template Preparation Kit for Genomic DNA DNAse/ RNAse free su MARKA Sigma Chemical Co., St Louis Missouri Sigma Chemical Co., St Louis Missouri Sigma Chemical Co., St Louis Missouri Biological Industries, Israel Biological Industries, Israel Biological Industries, Israel Biological Industries, Israel Biological Industries, Israel Zymo Research Roche Diagnostics, Almanya Biochrom Roche Applied Science, Mannheim, Germany Roche Diagnostics, Almanya Roche Diagnostics, Almanya Fermentas Olgu ve Hücre Hatları Ege Üniversitesi Nöroşirurji Anabilim Dalı nda beyin tümörü tanısı almış 98 olgu çalışıldı. Çalışmamıza katılmayı kabul etmiş, gönüllü onam formu imzalamış olgulardan tanı amaçlı rutin biyopsi uygulamaları esnasında veya operasyon ile rezeke edilen tümörden tümör örneği alındı. Tümör örneklerinin transferi 15ml lik falkon tüplerde bulunan serum içermeyen DMEM içerisinde operasyonun hemen akabinde gerçekleşti
101 Çalışmanın kontrollü ilerlemesi amacıyla, çeşitli hücre hatları kullanıldı. Pozitif kontrol olarak U87MG (Glioblastom, astrositom, derece IV), U-118MG (Glioblastoma, astrositom), LN-18 (beyin serebrum; sağ temporal lob, derece IV, glioblastoma; glioma), Daoy (beyin, serebellum desmoplastik serebellar medulloblastoma) hücre hatları ve negatif kontrol olarak WI-38 (Normal Akciğer fibroblastı) ve Hs 605.Sk (Deri; Fibroblastı, normal) hücre hatları kullanıldı Tümör Derece ve Evresinin Belirlenmesi Ege Üniversitesi Nöroşirurji Anabilim Dalı nda alınan 98 tümör dokusu, çalışılacak tümör örnekleri arasındaki heterojenitenin bertaraf edilmesi amacıyla 2 eşit parçaya ayrıldı. Aynı tümörden alınan parçalardan biri, Patoloji Anabilim Dalı nda eş zamanlı olarak tümörün tipi ve derecesini belirlemek üzere histopatolojik olarak değerlendirildi Tümör Hücre Kültürü Tümör dokusuna ait diğer parçasına 2 farklı hücre kültür yöntemi uygulandı. Birinci yöntemde doku mekanik olarak küçük parçalara ayrıldı ve bu eksplantlar kültüre edildi. (mekanik parçalama). Diğer yöntemde (enzimatik parçalama) ise, doku Kollegenaz Tip IV (Biochrom) ile enzimatik parçalanarak süspansiyon kültür yapıldı. In vitro koşullarda gerçekleştirilen hücre kültürü çalışmalarında kullanılan besi ortamı, hücrelerin in vitro koşullar altında yaşayabilmeleri, büyüyebilmeleri ve proliferasyonu için uygun olmalı ve gerekli maddeleri içermelidir. İnaktif serum, hücrelerin gereksinimlerini karşılamak amacıyla besi yerine mutlaka eklenmelidir.kültür ortamı olarak BIOAMF-1 (Biological Industiries), BIOAMF

102 supplement, 2mM L-glutamin, %1 Penisilin-Streptomisin kullanıldı. Hücrelerden yeterli üreme elde edilen kadar 37 o C de % 95 nem ve %5 CO 2 li etüvde inkübe edilmişlerdir. Hücrelerin proliferasyonu, pasajları ve takip işlemleri inverted mikroskop ile izlendi. Çalışmanın kapsamına uygun olan 100 olgunun 4 ayrı flaskte subkültürü yapıldı. Yüz olgunun flasklerinden total RNA, DNA izole edilmiş ve bir flaskteki çoğalan tümör hücreleri (Şekil 4) ise dondurulup, C de rezervuar olarak saklandı. Şekil 4: Beyin tümörlü olgunun hücre kültürünün inverted mikroskoptaki görüntüsü (300x) Negatif ve pozitif olarak kullanılan hücre hatlarının Hücre Kültürleri: U87MG (Glioblastom, astrositom), U-118MG (Glioblastoma), LN-18 (Glioblastoma), Daoy (Medulloblastoma) hücre hatları pozitif kontrol, WI
, BIOAMF-1 supplement, 2mM L-glutamin, %1 Penisilin-Streptomisin kullanıldı.")
103 (Akciğer fibroblastı) ve Hs 605.Sk (Deri; Fibroblastı) hücre hatları negatif kontrol olarak kullanıldı. Kültür ortamı olarak BIOAMF-1 (Biological Industiries), BIOAMF-1 supplement, 2mM L-glutamin, %1 Penisilin-Streptomisin kullanıldı. Hücrelerden yeterli üreme elde edilen kadar % 95 nem ve %5 CO 2 li etüvde inkübe edilmişlerdir. Hücrelerin proliferasyonu, pasajları ve takip işlemleri inverted mikroskop ile izlendi. Çalışmanın amacına uygun olarak her bir olgunun 4 ayrı flaskte subkültürü yapıldı. Flasklerinden total RNA, DNA izole edilmiş ve bir flaskteki çoğalan hücreler (Şekil 5) ise dondurulup C de rezervuar olarak saklandı. Çalışma kapsamında uygulanan yöntem Şekil 6 gösterilmiştir. (100x) Şekil 5: Kültüre edilen U-87 MG hücre hattının inverted mikroskop görüntüsü

104 Nöroşirurji Anabilim Dalı Tümör dokusunun alınması Tıbbi Biyoloji Anabilim Dalı Mekanik Hücre kültürü Süspansiyon Patoloji Anabilim Dalı Eş zamanlı tümör tanı ve derecesinin belirlenmesi DNA Promoter bölgelerin Gen kopya sayısının metilasyonunun kantitasyonu kantitasyonu Real-time Online PCR % metilasyon % Kayıp / kazanç Stok C RNA mrna kantitasyonu Real-time Online RT- PCR Gen ekspresyonlarının belirlenmesi Tanı Prognoz Beyin Tümörleri Tedavi Şekil 6: Çalışma kapsamında uygulanan yöntem Hücre Yedeklenmesi Tümör Hücrelerinin Dondurulması: 1) Flasklarda bulunan hücreler öncelikle tripsinizasyon işlemi ile yapışmış oldukları flask yüzeyinden kaldırıldı. 2) Hücreler 15 ml lik falkon tüplere alınarak 2000 rpm de 5 dakika süre ile santrifüj edildi. 3) Santrifüj sonrası, herbir falcon tüpe ait 1,5 ml lik kriyo tüpler hazırlandı
105 hazırlandı. 4) 9:1 oranında (%10 luk), serum: DMSO karışımından oluşan ortam 5) Tüplerde bulunan süpernatant kısım uzaklaştırılarak falkon tüpler buz üzerine alındı. Falkon tüpler de bulunan hücreler üzerine 750 µl serum: DMSO karışımı eklenerek resüspanse edildi. kaldırıldı. 6) Buz içerisinde bulunan kriyotüpler içerisine bu karışım aktarılarak -86 C ye Dondurulmuş Tümör Hücrelerinin Çözdürülmesi: 1) -86⁰C de kriyotüpler içerisinde bulunan %10 DMSO ile dondurulmuş hücreler 37⁰C ye alındı 2) DMSO nun hücre canlılığı üzerinde negatif etkisi bulunmaktadır. DMSO dan zarar görme sürecisini azaltmak amacıyla kriyotüp içerisine hızlı bir şekilde besi ortamı ilave edildi. Pasteur pipeti yardımıyla karıştırılarak hücreler 15 ml lik falkon tüp içerisine alındı. 3) Falkon tüp içerisine yaklaşık 10ml daha besi yeri ilave edilerek 1200 rpm de 5 dakika santrifuj edildi. 4) Santrifuj sonrası falkon içerisinde bulunan süpernatant uzaklaştırıldı ve hücreler üzerine tekrar (~10 ml) besi ortamı eklendi. 5) Tekrar 1200 rpm de 5 dakika santrifuj yapıldı
106 6) Santrifüj sonrası süpernatant uzaklaştırıldı ve çöken hücrelerin üzerine 5 ml besi yeri eklenerek 75 cm 2 lik flasklara alındı. bırakıldı. 7) Flasklar, 37⁰C deki %95 nem ve %5 CO 2 içeren etüve alınarak inkübasyona 48 saat sonra üremeye başlayan hücrelerin kullandıkları eski besi ortamı döküldü ve yerine 5 ml taze besi yeri ilave edildi. Yaklaşık 1 hafta sonra hücreler bütün flask yüzeyini kapladı. Hücrelerin canlılığını korumak ve devamlılığını sağlamak amacıyla pasaj yapıldı Tümör Hücrelerinin ve Hücre Hatlarının Hücrelerinin Pasajlanması 1) Tümör hücreleri ve hücre hatları hücreleri monolayer hücreler oldukları için hücrelerin pasajlanması için ilk olarak yapıştıkları yüzeyden ayrılmaları gerekir. Bu nedenle, flasklarda bulunan ortam döküldü ve flasklar bir kez serumsuz ortam kullanılarak yıkandı. 2) Daha sonra hücrelerin üzerine 2 ml tripsin ilave edildi. Bir süre bekletildikten sonra tripsin uzaklaştırılarak flasklar 5 dakika boyunca inkübatörde bekletildi. 3) İnkübatörden çıkan flasklar inverted mikroskopta incelendi ve tüm hücrelerin kalkıp kalkmadığı kontrol edildi. edildi. 4) Flaskda bulunan tüm hücreler kalktıktan sonra, 10 ml serumlu ortam ilave
 Hücrelerin yüzeye yapışarak üremeye devam etmeleri için flasklar inkübatöre kaldırılarak inkübasyona bırakıldı ve çoğalmaları sağlandı. 2.1.7.")
107 5) Flask içerisinde bulunan hücre-ortam karışımının 5 ml si başka bir flaska aktarılarak pasajlama işlemi tamamlanmış oldu. 6) Hücrelerin yüzeye yapışarak üremeye devam etmeleri için flasklar inkübatöre kaldırılarak inkübasyona bırakıldı ve çoğalmaları sağlandı Tümör Hücrelerinin ve Hücre hatlarının Canlılığının Belirlenmesi Hücre canlılığının belirlenmesinde tripan mavisi boyası testi uygulandı (Şekil 7). Tripan mavisi boyası canlı hücreler tarafından metabolize edildi ve hücre dışına atıldı. Bu nedenle, canlı hücreler beyaz renkte görüldü. Ölü hücreler ise, tripan mavisini metabolize edemedikleri için maviye boyandılar. Canlı hücre Ölü hücre Şekil 7: Eksklüzyon boya prensibi 1. Tümör hücreleri ve hücre hatları monolayer hücreler oldukları için ilk olarak tripsinizasyon işlemi yapıldı. 2. Tripsinizasyon işleminden sonra hücre süspansiyonundan 50 µl alınarak 96 gözlü plate lere aktarıldı ve 50 µl tripan mavisi boyası ile karıştırıldı. 3. Canlı ve ölü hücreler Neubauer lamda sayıldı
108 Total RNA İzolasyonu Gen ekspresyonunu değerlendirmek amacıyla tümör Hücrelerinden ve hücre hatlarından 1 er ml alınarak total RNA izolasyonu yapıldı. Total RNA izolasyonu için High Pure RNA Isolation Kit i (Roche Applied Science, Mannheim, Germany) kiti kullanıldı. Kültüre edilen hücrelerin trna izolasyonu için izlenen protokol: 1) Öncelikle kültürdeki hücre sayısı en fazla 1x10 6 olacak şekilde üreme elde edildi. 2) Hücre kültür kabındaki (flask) adesiv (yapışan) hücreleri kaldırmak amacıyla 500 µl Tripsin-EDTA solüsyonu ile muamele edildi. 3) 5 dakika CO 2 li etüvde inkübe edildi. 4) İnkübasyon sonrasında, flasklar inverted mikroskopta morfolojik olarak incelendi ve adesiv hücrelerin flask yüzeyinden kalkıp, yuvarlak hale geldikleri gözlemlendikten sonra üzerlerine 500µl serumlu medyum ilave edildi. 5) Tüm karışım 15 ml lik eppendorf tüplere aktarıldı. 6) Eppendorf tüpteki hücreler 2000 rpm de 5 dakika santrifüj edildi. 7) Santrifüj sonrasında ortam dökülüp, pellet üzerine 200µl soğuk (2-8 0 C) serum fizyolojik konuldu ve hücre pelleti resüspanse edildi. 8) Bunun üzerine, 400 µl Lizis bağlama tamponu eklendi. 9) Bu karışım pipetle iyice resüspanse edildikten sonra, cam lifli filtreli tüpe aktarıldı. 10) 10,000 rpm de 15 saniye santrifüj edildikten sonra, filtrenin altına geçen süpernatant atıldı
109 11) 90 µl DNAz inkübasyon tamponu + 10 µl DNAz dan oluşan karışım hazırlandı, sonra tamamı filtreli tüpe aktarıldı ve 15 dakika oda sıcaklığında inkübasyona bırakıldı. 12) İnkübasyon sonrasında, DNA parçalanır, filtrede sadece total RNA kalır. 13) İnkübasyonu tamamlanmış hücrelerin üzerine 500 µl Yıkama Tamponu I eklenip, 10,000 rpm de 15 saniye santrifüj yapılıp, süpernatant atıldı. 14) 500 µl Yıkama Tamponu II eklenip, 10,000 rpm de 15 saniye santrifüj yapılıp, süpernatant atıldı. 15) Hücrelerin üzerine 200 µl Yıkama Tamponu II eklenip, rpm de 2 dakika santrifüj yapıldı. 16) Filtre, yeni bir Eppendorf tüpe aktarıldı. 50 µl elüsyon tamponu eklenip, 10,000 rpm de 1 dakika santrifüj yapıldı. 17) Santrifüj sonrası 1,5 ml tüpte kalan total RNA dır. Elde edilen total RNA, komplementer DNA elde edilecek süreye kadar C buzdolabında saklandı Revers Transkriptaz RT-PCR cdna Sentezi Total RNA ları elde edilen tümör örneklerinden 1-5 µg/ml olacak şekilde Transcriptor First Strand cdna Synthesis Kit (Roche Diagnostics) kullanılarak cdna sentezleri 2 aşamada gerçekleştirildi (Tablo 3-4)
110 RNAz ve DNAz içermeyen PCR tüplerindeki total revers transkriptaz reaksiyon volümü 20µl olacak şekilde işleme başlandı. Tablo 3: Komplementer DNA analizinin ilk basamağı Ajan Hacim Son miktar Su 1µl Total RNA 10µl 1µg (10ng-5µg) Anchored-oligo (dt) 18 primeri 1µl 2,5 µm (50 pmol/µl) Son hacim 13µl Buz üzerinde 65 O C de 10 dakika inkübasyon (Bu basamak RNA nın sekonder yapılarının denatürasyonu için) Tablo 4: cdna analizinin ikinci basamağı Ajan Hacim Son miktar Transkriptör RT Reaksiyon Tamponu 4 µl 1X Koruyucu RNAz İnhibitörü (40U/µl) 0,5 µl 20 U Deoksinukleotid karışımı 2 µl Her biri için 1mM (her biri için 10mM) Transkriptör Revers Transkriptaz 0,5 µl 10 U Son hacim 20 µl 55 0 C de 30 dakika inkübasyondan sonra reaksiyon tübündeki elde edilen cdna uzun dönem saklanabilmek için C ye kaldırıldı Real-time Online Revers Transkriptaz RT-PCR ve Taqman Protokolü Real-time Online RT-PCR gen ekspresyon ürününü kantite etme amacıyla kullanılan moleküler bir metottur. PCR reaksiyonlarında sıcaklık döngülerini sağlamak için kullanılan cihazların (thermocycler), hassas ölçüm aletleri ile birleştirilmesi sonucu gerçek zamanlı PCR yöntemi geliştirildi. Bu yöntem sayesinde,
111 DNA ve RNA örnekleri kalitatif ve kantitatif olarak kısa sürede analiz edilebilmekte, çok sayıda örnek son derece düşük kontaminasyon riskiyle güvenle çalışılabilmektedir. Gerçek zamanlı PCR / RT-PCR da ürünlerin analizi reaksiyon esnasında yapılmaktadır. Bu nedenle, agaroz jel elektroforezi, PCR ürününün mor ötesi ışık altında görüntülenmesi gibi işlemlerin uygulanmasına gerek kalmamaktadır. Real-time Online PCR / RT-PCR ürünlerinin kalitatif ve kantitatif analizlerinde diziye özgün problardan yararlanılmaktadır. Böylece sonuçlar anında alınmakta, kontaminasyon riski azalarak tüm işlemler sıcaklık döngüleri başlayınca otomatik olarak devam etmektedir. Ekspresyon analizinde kullanılan primer ve problar: Tümör Süpresör ve Onkogen Gen Ekspresyon Analizlerinde Kullanılan Primer ve Problar İlgili Primer ve problar web tabanlı ProbeFinder Software i ( kullanılarak dizayn edildi (Tablo 5-6). Tablo 5: Tümör süpresörler için belirlenen ilgili primer ve problar Gen İleri Primer Geri Primer Prob DMBT1 gctcaggaaaccatctatcgac gaagcctccgcaggaatagt ctgctcct MGMT gcccctagaacgctttgc ttgtccatttttccaagtacca ccgccgcc PIKE aacaccatccaagactgaagg aggattttagtttccacattttgc cttcagcc PTEN ggggaagtaaggaccagagac tccagatgattctttaacaggtagc ttcccagt RB1 cagaataatcacactgcagcagata cacgcgtagttgaaccttttt caggagaa RUNX3 tcagcaccacaagccactt aatgggttcagttccgaggt gcagccag TP53 ccccagccaaagaagaaac aacatctcgaagcgctcac ggatggag GAPDH gaaggtgaaggtcggagtc gaagatggtgatgggatttc caagcttcccgttctcagcc
112 Tablo 6: Onkogenler için belirlenen ilgili primer ve problar Gen İleri Primer Geri Primer Prob COX-2 tcacgcatcagtttttcaaga tcaccgtaaatatgatttaagtccac gggctggg EGFR cagccacccatatgtaccatc aactttgggcgactatctgc gctggatg GSTT1 acggggacttcaccttgac gaccttatatttgcgcgtcag tcctgctc htert gccttcaagagccacgtc ccacgaactgtcgcatgt ctccagcc Mdm2 ccatgatctacaggaacttggtagta tcactcacagatgtacctgagtcc cagcagga OLIG2 tctctcttttctcctcctctcct atagtcgtcgcagctttcg ggaaggag PIK3CA cacgagatcctctctctgaaatc ggtagaatttcggggatagttaca caggagaa TaqMan PCR Protokolü PCR, Denatürasyon, Amplifikasyon ve Soğutma aşamalarını içermektedir. 1.Denatürasyon Döngü Programı Verileri Değer Döngü 1 Analiz Modu - Hedef Dereceleri Segment 1 Hedef Derece ( C) 95 İnkübasyon zamanı (sn) 600 Sıcaklık Transisyon Oranı ( C/sn) Amplifikasyon Döngü Programı Verileri Değer Döngü 40 Analiz Modu Kantitasyon Hedef Dereceleri Segment 1 Segment 2 Hedef Derece ( C) İnkübasyon zamanı (sn) Sıcaklık Transisyon Oranı ( C/san) 20,0 20,0 İkinci Hedef Derecesi ( C) 0 0 Basamak derecesi ( o C) 0,0 0,0 Döngüler 0 0 Kazanç modu - Tek
113 3.Soğuma Program Verileri Değer Döngü sayısı 1 Analiz Modu - Hedef Dereceleri Segment 1 Hedef Derece ( C) 40 İnkübasyon Zamanı 30 Sıcaklık Transisyon Oranı ( C/san) 20, Gen Ekspresyonun Değerlendirilmesi Gerçek zamanlı RT-PCR da ürünlerin analizi reaksiyon esnasında yapılmaktadır. Bu nedenle, agaroz jel elektroforezi, DNA bantlarının mor ötesi ışık altında görüntülenmesi gibi işlemlerin uygulanmasına gerek kalmamaktadır. Gerçek zamanlı PCR ürünlerinin kalitatif ve kantitatif analizlerinde, diziye özgün olmayan flüoresan boyalardan ya da diziye özgün problardan yararlanılmaktadır. Böylece sonuçlar anında alınmakta, kontaminasyon riski azalarak tüm işlemler sıcaklık döngüleri başlayınca otomatik olarak devam etmektedir. Tıbbi Biyoloji Anabilim Dalımızda aktif olarak kullandığımız bu cihaz ile çalışmamızda aldığımız sonuçların güvenirliliği oldukça yüksektir. LightCycler, gerçek zamanlı (real-time) bir PCR cihazıdır. Bu cihazda, amplifikasyon ürünlerinin artışı eş zamanlı olarak takip edilebilmektedir. Oldukça hızlı ısıtma ve soğutma kapasitesi sayesinde, PCR döngüsü, dakika içinde yapılabilmektedir. Sistemde, erime eğrisi analizi sayesinde, diziye özgül hibridizasyon problarının tek zincirli DNA ya hibridizasyonu izlenebilmektedir. Araştırılan gen bölgesi, o bölgeye özgül iki primer ile amplifiye edildikten sonra, oluşan amplikon, bir sonraki döngünün annealing aşamasında
114 özgül hibridizasyon probları ile tespit edilmektedir. Oligo 1, 3 ucundan fluorescein (donör boya) ile oligo 2 ise, 5 ucundan LCRed640 (alıcı boya) ile işaretlendi. Annealing aşamasında iki oligonükleotid, DNA fragmanına baş kuyruk pozisyonunda bağlanmaktadır. Oligonükleotidler bu pozisyonda hibridize olduklarında iki florasan boya birbirine çok yaklaşmış olmaktadır. Fluorescein ile işaretli 1. oligo, LC cihazının ışık kaynağından yayılan 470 nm deki mavi ışık ile uyarılmakta ve uzun dalga boyunda (530 nm) yeşil florasan ışık yaymaktadır. İki probun birbirine çok yakın bulundukları annealing aşamasında 530 nm de yayılan enerji, ikinci hibridizasyon probuna aktarılarak onu uyarmakta ve farklı bir dalga boyunda (640 nm) kırmızı florasan ışık yayılmasına neden olmaktadır. İki prob arasındaki bu enerji transferi, Floresan Rezonans Enerji Transferi yani FRET olarak tanımlanmaktadır. PCR dan sonra herhangi ek bir manuel işleme ihtiyaç yoktur ve buna bağlı olarak, kontaminasyon riski de en aza indirgendi. İki µl cdna, Light Cycler PCR Master Mix ten 7µl kullanılarak, PCR ile amplifiye edilir. PCR sırasında, mrna primerler ile amplifiye edildi. Taqman sisteminde, 5 ve 3 uçlarından florokrom maddelerle işaretli prob kullanılmaktadır. Probun 5 ucunda raportör florokrom (6-karboksifloresein=6-FAM), 3 ucunda ise baskılayıcı florokrom (6-karboksi-tetrametil-rodamin=TAMRA) bulunmaktadır. Prob tek sarmal hale getirilen hedef molekül üzerinde, primerlerin bağlanma bölgesinin arasında kalan yere bağlanmaktadır. Prob-hedef molekül arasındaki hibridizasyon devam ettiği sürece raportör florokrom maddenin sinyal oluşturması, 3 uçtaki baskılayıcı florokrom tarafından engellenmektedir. Primerlerin hedef nükleik aside bağlanmasını takiben başlatılan primer uzaması probun bağlandığı noktaya kadar
115 geldiğinde, sentezin devam edebilmesi için Taq DNA Polimeraz enzimi 5 3 ekzonükleaz aktivitesini kullanarak probu 5 uçtan yıkmaya başlar. Böylece rapörtor florokrom serbest hale geçer ve sinyal oluşturur. Her siklusta üretilen amplikon miktarına paralel olarak sinyal şiddeti de artmaktadır. Kalıp DNA ya hibridizasyondan sonra yansıtılan flüoresan Light Cycler cihazı ile ölçülür. Ayrı bir PCR da yapılan aynı cdna preperasyonu kullanılarak, gliseraldehit-3-fosfat dehidrogenaz (GAPDH) cdna sı house keeping gen olarak kullanılır. Bunun ürünü RNA kontrolü ve rölatif kantitasyon için kullanılır. Her bir örneğin kantitasyonu, örneğin flüoresan rezonans enerjisi Light Cycler Kantitasyon Software ı (LCQuant, Roche Diagnostics) kullanılarak ölçülür. İncelenecek örnekte, genlerin mrna miktarının ölçülmesi için rölatif kantitasyon kullanılır. Amplifiye edilen DNA nın kantitasyon analizi, kantitasyon software (Relative Quantification software Roche Diagnostics GmbH, Mannheim, Germany) ile hesaplanmaktadır. PCR sırasında elde edilen amplikonlar, doğrudan LightCycler cihazında, logaritmik artışa geçtikleri döngü sayısına göre değerlendirilmektedir. Bunun için önce konsantrasyonları bilinen GAPDH ın 7 farklı standardı ile (10 8,10 7, 10 6, 10 5, ve 10 2 ) bir standart amplifikasyon eğrisi oluşturulur. Çalışılan örnekteki geçiş noktasına (CP) göre cdna nın rölatif miktarı kantitasyon yazılımı tarafından belirlenmektedir (Şekil 8). Hedef gen kopya sayısının logaritmasının referans (housekeeping) gen kopyası logaritmasına oranı ilgili genin ekspresyonunun rölatif oranı (RR) ifade etmektedir. Log (Hedef gen kopya sayısı) RR= Log (Referans gen kopya sayısı)


116 Cp Cp A b Şekil 8: (a) Hedef gen, (b) Housekeeping gen geçiş noktası Gen Kopya Sayısı DNA İzolasyonu Tümör süpresör genlerin, gen kopya sayılarının analizi için kültüre edilen hücrelerden High Pure PCR Template Preparation Kit for Genomik DNA (Roche Diagnostics, Almanya) kullanılarak kit manueline uygun olarak DNA izolasyonu gerçekleştirildi. Hücreler 1 er ml 1.5 ml lik steril tüplere alınıp, 1200 RPM de 5 dakika santrifüj ile çöktürüldükten sonra üzerine ve 200 µl bağlama tamponu ve 40 µl Proteinaz K pipetlendi. Karıştırıldı ve 72 C de 10 dakika inkübe edildi. İnkübasyonun sonunda karışıma 100 ml izopropanol eklendi ve pipetaj ile iyice karıştırıldı. Karışım bir collection- toplama tübünün içine yerleştirilmiş filtre tübünün içine pipetlendi, rpm de 1 dakika santrifüj edildi. Altta biriken sıvı
DİFÜZ GLİAL TÜMÖRLER
 DİFÜZ GLİAL TÜMÖRLER DSÖ 2016 da erişkin glial tümörler açısından sizce en önemli değişiklik ne olmuştur? Curr Opin Oncol 2016 Nov;28(6):494-501. Diffuz astrositik ve oligodentroglial tümörler aynı grup
DİFÜZ GLİAL TÜMÖRLER DSÖ 2016 da erişkin glial tümörler açısından sizce en önemli değişiklik ne olmuştur? Curr Opin Oncol 2016 Nov;28(6):494-501. Diffuz astrositik ve oligodentroglial tümörler aynı grup
SANTRAL SİNİR SİSTEMİ TÜMÖRLERİ
 SANTRAL SİNİR SİSTEMİ TÜMÖRLERİ Tümörlerin yerleşim yerleri ile ilgili kullanılan terimler İntraaksiyel, ekstraaksiyel İntrakranyal Supratentoryal, infratentoryal Orta hat (sella, 3.ventrikül, pineal,
SANTRAL SİNİR SİSTEMİ TÜMÖRLERİ Tümörlerin yerleşim yerleri ile ilgili kullanılan terimler İntraaksiyel, ekstraaksiyel İntrakranyal Supratentoryal, infratentoryal Orta hat (sella, 3.ventrikül, pineal,
Beyin tümörlerinde genel olarak kabul görmüş sınıflandırma WHO (World. Health Organization) tarafından yapılmış olan ve periyodik olarak yenilenen
 tarafından yapılmış olan ve periyodik olarak yenilenen") Beyin Tümörleri Dr. Fazıl Gelal Dr. Kamil Karaali Beyin tümörlerinde genel olarak kabul görmüş sınıflandırma WHO (World Health Organization) tarafından yapılmış olan ve periyodik olarak yenilenen sınıflandırmadır.
Beyin Tümörleri Dr. Fazıl Gelal Dr. Kamil Karaali Beyin tümörlerinde genel olarak kabul görmüş sınıflandırma WHO (World Health Organization) tarafından yapılmış olan ve periyodik olarak yenilenen sınıflandırmadır.
İntrakranial. Gaziantep Üniversitesi Tıp Fakültesi Nöroşirürji Kliniği. Prof.Dr.Abdülvahap GÖK
 İntrakranial Tümörler Gaziantep Üniversitesi Tıp Fakültesi Nöroşirürji Kliniği Prof.Dr.Abdülvahap GÖK TANIM -Primer beyin tümörleri santral sinir sistemi dokusundan gelişir ve intrakranial tümörlerin 1/2
İntrakranial Tümörler Gaziantep Üniversitesi Tıp Fakültesi Nöroşirürji Kliniği Prof.Dr.Abdülvahap GÖK TANIM -Primer beyin tümörleri santral sinir sistemi dokusundan gelişir ve intrakranial tümörlerin 1/2
Tanımlar. Grade: Yayılma eğilimi, Büyüme hızı, Normal hücrelere benzerlik. Grade I-IV.
 Beyin Tümörleri Tanımlar Primer/ Sekonder: %50-%50 İntra/Ekstraaksiyal Grade: Yayılma eğilimi, Büyüme hızı, Normal hücrelere benzerlik. Grade I-IV. Lokalizasyona göre sınıflama Supra/İnfratentoryel Erişkin:
Beyin Tümörleri Tanımlar Primer/ Sekonder: %50-%50 İntra/Ekstraaksiyal Grade: Yayılma eğilimi, Büyüme hızı, Normal hücrelere benzerlik. Grade I-IV. Lokalizasyona göre sınıflama Supra/İnfratentoryel Erişkin:
İNTRAKRANYAL TÜMÖRLER. CEM CALLI Dept. Of Radiology, Section of Radiology, Ege University, Izmir, Turkey
 İNTRAKRANYAL TÜMÖRLER CEM CALLI Dept. Of Radiology, Section of Radiology, Ege University, Izmir, Turkey İNTRAKRANYAL TÜMÖRLER SINIFLAMA: * Histolojik (WHO 2000) * Yaş (pediatrik, erişkin) * Yerleşim(intra/extra-aksiyel,
İNTRAKRANYAL TÜMÖRLER CEM CALLI Dept. Of Radiology, Section of Radiology, Ege University, Izmir, Turkey İNTRAKRANYAL TÜMÖRLER SINIFLAMA: * Histolojik (WHO 2000) * Yaş (pediatrik, erişkin) * Yerleşim(intra/extra-aksiyel,
BEYİN TÜMÖRLERİ: RİSK FAKTÖRLERİ. 1. Ailesel hastalıklarla ilişkisi
 BEYİN TÜMÖRLERİ: Beyin tümörleri çocukluk çağında görülen en sık solid tümördür. Bu yaş grubundaki çocuklarda görülen kanserlerin %20 sini oluşturur. 15 yaşın altında görülen malign hst. içinde lösemilerden
BEYİN TÜMÖRLERİ: Beyin tümörleri çocukluk çağında görülen en sık solid tümördür. Bu yaş grubundaki çocuklarda görülen kanserlerin %20 sini oluşturur. 15 yaşın altında görülen malign hst. içinde lösemilerden
GLİAL TÜMÖRLERDE AYIRICI TANIDA KULLANILAN İMMUNOHİSTOKİMYASAL IDH-1, ATRX, p53 ve FISH 1p19q KODELESYONU
 GLİAL TÜMÖRLERDE AYIRICI TANIDA KULLANILAN İMMUNOHİSTOKİMYASAL IDH-1, ATRX, p53 ve FISH 1p19q KODELESYONU DR.NESLİHAN KAYA,ŞEBNEM BATUR,NİL ÇOMUNOĞLU,BÜGE ÖZ İ.Ü.CERRAHPAŞA TIP FAKÜLTESİ PATOLOJİ ANABİLİM
GLİAL TÜMÖRLERDE AYIRICI TANIDA KULLANILAN İMMUNOHİSTOKİMYASAL IDH-1, ATRX, p53 ve FISH 1p19q KODELESYONU DR.NESLİHAN KAYA,ŞEBNEM BATUR,NİL ÇOMUNOĞLU,BÜGE ÖZ İ.Ü.CERRAHPAŞA TIP FAKÜLTESİ PATOLOJİ ANABİLİM
Akciğer Karsinomlarının Histopatolojisi
 Akciğer Karsinomlarının Histopatolojisi Journal of Clinical and Analytical Medicine Göğüs Cerrahisi Akciğer karsinomlarının gelişiminde preinvaziv epitelyal lezyonlar; Akciğer karsinomlarının gelişiminde
Akciğer Karsinomlarının Histopatolojisi Journal of Clinical and Analytical Medicine Göğüs Cerrahisi Akciğer karsinomlarının gelişiminde preinvaziv epitelyal lezyonlar; Akciğer karsinomlarının gelişiminde
Cerrahi Patologlar için Nöroradyoloji
 Cerrahi Patologlar için Nöroradyoloji Doç. Dr. Önder Öngürü 27. Ulusal Patoloji Kongresi Antalya, 15 Kasım 2017 Hedef 1. Sık tümörlerin alışılageldik radyolojik görünümlerini tanıyabilmek 2. Mutad dışı
Cerrahi Patologlar için Nöroradyoloji Doç. Dr. Önder Öngürü 27. Ulusal Patoloji Kongresi Antalya, 15 Kasım 2017 Hedef 1. Sık tümörlerin alışılageldik radyolojik görünümlerini tanıyabilmek 2. Mutad dışı
DİĞER EMBRİYONAL TÜMÖRLER II.BÖLÜM. DR. NİL ÇOMUNOĞLU İ.Ü. Cerrahpaşa Tıp Fakültesi Patoloji Anabilim Dalı
 DİĞER EMBRİYONAL TÜMÖRLER II.BÖLÜM DR. NİL ÇOMUNOĞLU İ.Ü. Cerrahpaşa Tıp Fakültesi Patoloji Anabilim Dalı 2007 2016 DİĞER EMBRİYONAL TÜMÖRLER-WHO GRADE IV-2016 ATİPİK/TERATOİD RABDOİD TÜMÖR RABDOİD ÖZELLİKLİ
DİĞER EMBRİYONAL TÜMÖRLER II.BÖLÜM DR. NİL ÇOMUNOĞLU İ.Ü. Cerrahpaşa Tıp Fakültesi Patoloji Anabilim Dalı 2007 2016 DİĞER EMBRİYONAL TÜMÖRLER-WHO GRADE IV-2016 ATİPİK/TERATOİD RABDOİD TÜMÖR RABDOİD ÖZELLİKLİ
SAĞ VE SOL KOLON YERLEŞİMLİ TÜMÖRLER: AYNI ORGANDA FARKLI PATOLOJİK BULGULAR VE MİKROSATELLİT İNSTABİLİTE DURUMU
 SAĞ VE SOL KOLON YERLEŞİMLİ TÜMÖRLER: AYNI ORGANDA FARKLI PATOLOJİK BULGULAR VE MİKROSATELLİT İNSTABİLİTE DURUMU Ezgi Işıl Turhan 1, Nesrin Uğraş 1, Ömer Yerci 1, Seçil Ak 2, Berrin Tunca 2, Ersin Öztürk
SAĞ VE SOL KOLON YERLEŞİMLİ TÜMÖRLER: AYNI ORGANDA FARKLI PATOLOJİK BULGULAR VE MİKROSATELLİT İNSTABİLİTE DURUMU Ezgi Işıl Turhan 1, Nesrin Uğraş 1, Ömer Yerci 1, Seçil Ak 2, Berrin Tunca 2, Ersin Öztürk
Türk Nöroşirurji Derneği. Temel Nöroşirurji Kursları Dönem 4, 1. Kurs. Program
 Temel Nöroşirurji Kursları Dönem 4, 1. Kurs 16 Mart 2016, Çarşamba OTELE GİRİŞ 16:00 18:00 OTURUM Başkan: Selçuk Palaoğlu GİRİŞ Nöroonkolojik cerrahi; Beyin: Genel kavramlar, hasta takip sınıflamalar ve
Temel Nöroşirurji Kursları Dönem 4, 1. Kurs 16 Mart 2016, Çarşamba OTELE GİRİŞ 16:00 18:00 OTURUM Başkan: Selçuk Palaoğlu GİRİŞ Nöroonkolojik cerrahi; Beyin: Genel kavramlar, hasta takip sınıflamalar ve
2016 DSÖ beyin tümörleri sınıflaması
 2016 DSÖ beyin tümörleri sınıflaması Neler değişti? Neler değişti Dr Büge Öz İÜ Cerrahpaşa Tıp Fakültesi Patoloji AD 1979 1993 2000 2007 Morfolojiye dayalı sınıflama Molekülere giriş 2016 Morfoloji + Moleküler
2016 DSÖ beyin tümörleri sınıflaması Neler değişti? Neler değişti Dr Büge Öz İÜ Cerrahpaşa Tıp Fakültesi Patoloji AD 1979 1993 2000 2007 Morfolojiye dayalı sınıflama Molekülere giriş 2016 Morfoloji + Moleküler
MİDE KANSERİNDE APOPİTOZİSİN BİYOLOJİK BELİRTEÇLERİNİN PROGNOSTİK ÖNEMİ
 MİDE KANSERİNDE APOPİTOZİSİN BİYOLOJİK BELİRTEÇLERİNİN PROGNOSTİK ÖNEMİ Cem Sezer 1, Mustafa Yıldırım 2, Mustafa Yıldız 2, Arsenal Sezgin Alikanoğlu 1,Utku Dönem Dilli 1, Sevil Göktaş 1, Nurullah Bülbüller
MİDE KANSERİNDE APOPİTOZİSİN BİYOLOJİK BELİRTEÇLERİNİN PROGNOSTİK ÖNEMİ Cem Sezer 1, Mustafa Yıldırım 2, Mustafa Yıldız 2, Arsenal Sezgin Alikanoğlu 1,Utku Dönem Dilli 1, Sevil Göktaş 1, Nurullah Bülbüller
Beyin Omurilik ve Sinir Tümörlerinin Cerrahisi. (Nöro-Onkolojik Cerrahi)
") Beyin Omurilik ve Sinir Tümörlerinin Cerrahisi (Nöro-Onkolojik Cerrahi) BR.HLİ.018 Sinir sisteminin (Beyin, omurilik ve sinirlerin) tümörleri, sinir dokusunda bulunan çeşitli hücrelerden kaynaklanan ya
Beyin Omurilik ve Sinir Tümörlerinin Cerrahisi (Nöro-Onkolojik Cerrahi) BR.HLİ.018 Sinir sisteminin (Beyin, omurilik ve sinirlerin) tümörleri, sinir dokusunda bulunan çeşitli hücrelerden kaynaklanan ya
En Etkili Kemoterapi İlacı Seçimine Yardımcı Olan Moleküler Genetik Test
 En Etkili Kemoterapi İlacı Seçimine Yardımcı Olan Moleküler Genetik Test Yeni Nesil DNA Dizileme (NGS), İmmünHistoKimya (IHC) ile Hastanızın Kanser Tipinin ve Kemoterapi İlacının Belirlenmesi Kanser Tanı
En Etkili Kemoterapi İlacı Seçimine Yardımcı Olan Moleküler Genetik Test Yeni Nesil DNA Dizileme (NGS), İmmünHistoKimya (IHC) ile Hastanızın Kanser Tipinin ve Kemoterapi İlacının Belirlenmesi Kanser Tanı
MEME KANSERİ KÖK HÜCRELERİNİN GEN EKSPRESYON PROFİLİ
 MEME KANSERİ KÖK HÜCRELERİNİN GEN EKSPRESYON PROFİLİ Sait Murat Doğan, A. Pınar Erçetin, Zekiye Altun, Duygu Dursun, Safiye Aktaş Dokuz Eylül Üniversitesi Onkoloji Enstitüsü, İzmir Slayt 1 / 14 Meme Kanseri
MEME KANSERİ KÖK HÜCRELERİNİN GEN EKSPRESYON PROFİLİ Sait Murat Doğan, A. Pınar Erçetin, Zekiye Altun, Duygu Dursun, Safiye Aktaş Dokuz Eylül Üniversitesi Onkoloji Enstitüsü, İzmir Slayt 1 / 14 Meme Kanseri
Dr. Figen Söylemezoğlu
 Dr. Figen Söylemezoğlu Konsültasyon Tanıya ulaşmada zorluk Klinik ile uyumsuzluk Tedavinin bir başka kurumda devam etmesi Hasta ya da klinisyenin ikinci bir görüş istemesi durumunda Konsültasyon Bu uygulamanın
Dr. Figen Söylemezoğlu Konsültasyon Tanıya ulaşmada zorluk Klinik ile uyumsuzluk Tedavinin bir başka kurumda devam etmesi Hasta ya da klinisyenin ikinci bir görüş istemesi durumunda Konsültasyon Bu uygulamanın
Beyin Tümörleri. Prof. Dr.Murat Söker
 Beyin Tümörleri Prof. Dr.Murat Söker Beyin tümörleri çocukluk çağında görülen en sık solid tümör Bu yaş grubundaki çocuklarda görülen kanserlerin %20 15 yaşın altında görülen malign hst. içinde lösemilerden
Beyin Tümörleri Prof. Dr.Murat Söker Beyin tümörleri çocukluk çağında görülen en sık solid tümör Bu yaş grubundaki çocuklarda görülen kanserlerin %20 15 yaşın altında görülen malign hst. içinde lösemilerden
İntradural Spinal Tümörler. Dr. Fuldem Yıldırım Dönmez Başkent Üniversitesi Radyoloji Anabilim Dalı, Ankara
 İntradural Spinal Tümörler Dr. Fuldem Yıldırım Dönmez Başkent Üniversitesi Radyoloji Anabilim Dalı, Ankara Intraspinal tümörler spinal kord, filum terminale, sinir kökleri, meninksler, intraspinal damarlar,
İntradural Spinal Tümörler Dr. Fuldem Yıldırım Dönmez Başkent Üniversitesi Radyoloji Anabilim Dalı, Ankara Intraspinal tümörler spinal kord, filum terminale, sinir kökleri, meninksler, intraspinal damarlar,
NAZOFARENKS KARSİNOMUNDA CLAUDIN 1, 4 VE 7 EKSPRESYON PATERNİ VE PROGNOSTİK ÖNEMİ
 NAZOFARENKS KARSİNOMUNDA CLAUDIN 1, 4 VE 7 EKSPRESYON PATERNİ VE PROGNOSTİK ÖNEMİ Dinç Süren 1, Mustafa Yıldırım 2, Vildan Kaya 3, Ruksan Elal 1, Ömer Tarık Selçuk 4, Üstün Osma 4, Mustafa Yıldız 5, Cem
NAZOFARENKS KARSİNOMUNDA CLAUDIN 1, 4 VE 7 EKSPRESYON PATERNİ VE PROGNOSTİK ÖNEMİ Dinç Süren 1, Mustafa Yıldırım 2, Vildan Kaya 3, Ruksan Elal 1, Ömer Tarık Selçuk 4, Üstün Osma 4, Mustafa Yıldız 5, Cem
Sinir Sistemi Biyopsi ve Rezeksiyonları Patolojik Değerlendirme Standartları Ve Kılavuzu. (Nöropatoloji Çalışma Grubu)
") Sinir Sistemi Biyopsi ve Rezeksiyonları Patolojik Değerlendirme Standartları Ve Kılavuzu (Nöropatoloji Çalışma Grubu) 2010 Đçindekiler Genel Özellikler 2 Beyin/Spinal kord:biyopsi, rezeksiyon, stereotaktik
Sinir Sistemi Biyopsi ve Rezeksiyonları Patolojik Değerlendirme Standartları Ve Kılavuzu (Nöropatoloji Çalışma Grubu) 2010 Đçindekiler Genel Özellikler 2 Beyin/Spinal kord:biyopsi, rezeksiyon, stereotaktik
TTOD MEME KANSERİ GÜNCELLEME KURSU 13-14 HAZİRAN 2015 İSTANBUL 08:25-08:30 Açılış 08:00-08:30 Pratiği değiştiren çalışmalar. (salonda kahvaltı ile)
") TTOD MEME KANSERİ GÜNCELLEME KURSU 13-14 HAZİRAN 2015 İSTANBUL 08:25-08:30 Açılış 08:00-08:30 Pratiği değiştiren çalışmalar. (salonda kahvaltı ile) 1. Gün 1. Oturum: Meme kanserine giriş, Patoloji ve Alt
TTOD MEME KANSERİ GÜNCELLEME KURSU 13-14 HAZİRAN 2015 İSTANBUL 08:25-08:30 Açılış 08:00-08:30 Pratiği değiştiren çalışmalar. (salonda kahvaltı ile) 1. Gün 1. Oturum: Meme kanserine giriş, Patoloji ve Alt
KANSER EPİDEMİYOLOJİSİ VE KARSİNOGENEZ
 KANSER EPİDEMİYOLOJİSİ VE KARSİNOGENEZ Gökhan Erdem GATA Tıbbi Onkoloji BD 19 Mart 2014 5. Türk Tıbbi Onkoloji Kongresi, 19-23 Mart 2014, Antalya EPİDEMİYOLOJİ Epidemiyoloji, sağlık olaylarının görünme
KANSER EPİDEMİYOLOJİSİ VE KARSİNOGENEZ Gökhan Erdem GATA Tıbbi Onkoloji BD 19 Mart 2014 5. Türk Tıbbi Onkoloji Kongresi, 19-23 Mart 2014, Antalya EPİDEMİYOLOJİ Epidemiyoloji, sağlık olaylarının görünme
Sinir Kılıfı Tümörleri. Doç. Dr. Halil KIYICI 2016
 Sinir Kılıfı Tümörleri Doç. Dr. Halil KIYICI 2016 Sinir Sistemi Merkezi (santral) sinir sistemi (MSS): beyin ve omurilikten oluşur kafatası - omurga kemikleri ve kemik altındaki pia örtüsüyle kaplıdır
Sinir Kılıfı Tümörleri Doç. Dr. Halil KIYICI 2016 Sinir Sistemi Merkezi (santral) sinir sistemi (MSS): beyin ve omurilikten oluşur kafatası - omurga kemikleri ve kemik altındaki pia örtüsüyle kaplıdır
MESANE TÜMÖRLERİNİN DOĞAL SEYRİ
 MESANE TÜMÖRLERİNİN DOĞAL SEYRİ ve MOLEKÜLER PROGNOSTİK FAKTÖRLER Prof. Dr. Levent Türkeri Üroloji Anabilim Dalı Marmara Üniversitesi Tıp Fakültesi Mesane Tümörü (Transizyonel Hücreli Karsinom) Yüzeyel
MESANE TÜMÖRLERİNİN DOĞAL SEYRİ ve MOLEKÜLER PROGNOSTİK FAKTÖRLER Prof. Dr. Levent Türkeri Üroloji Anabilim Dalı Marmara Üniversitesi Tıp Fakültesi Mesane Tümörü (Transizyonel Hücreli Karsinom) Yüzeyel
SANTRAL SİNİR SİSTEMİ TÜMÖRLERİ VE GENETİK. Yrd. Doç. Dr. Ünal ULUCA
 SANTRAL SİNİR SİSTEMİ TÜMÖRLERİ VE GENETİK Yrd. Doç. Dr. Ünal ULUCA Primer santral sinir sistemi (SSS) tümörleri heterojen bir grup hastalıktır. Çocukluk çağında ve adolesan dönemde görülen ikinci en sık
SANTRAL SİNİR SİSTEMİ TÜMÖRLERİ VE GENETİK Yrd. Doç. Dr. Ünal ULUCA Primer santral sinir sistemi (SSS) tümörleri heterojen bir grup hastalıktır. Çocukluk çağında ve adolesan dönemde görülen ikinci en sık
İKİNCİL KANSERLER. Dr Aziz Yazar Acıbadem Üniversitesi Tıp Fakültesi İç Hastalıkları AD. Tıbbi Onkoloji BD. 23 Mart 2014, Antalya
 İKİNCİL KANSERLER Dr Aziz Yazar Acıbadem Üniversitesi Tıp Fakültesi İç Hastalıkları AD. Tıbbi Onkoloji BD. 23 Mart 2014, Antalya Tanım Kanser tedavisi almış veya kanser öyküsü olan bir hastada histopatolojik
İKİNCİL KANSERLER Dr Aziz Yazar Acıbadem Üniversitesi Tıp Fakültesi İç Hastalıkları AD. Tıbbi Onkoloji BD. 23 Mart 2014, Antalya Tanım Kanser tedavisi almış veya kanser öyküsü olan bir hastada histopatolojik
Tanı: Metastatik hastalık için patognomonik bir radyolojik. Tek veya muitipl nodüller iyi sınırlı veya difüz. Göğüs Cerrahisi Hasan Çaylak
 Journal of Clinical and Analytical Medicine Göğüs Cerrahisi Hasan Çaylak Göğüs Cerrahisi Metastatik Akciğer Tümörleri Giriş İzole akciğer metastazlarına tedavi edilemez gözüyle bakılmamalıdır Tümör tipine
Journal of Clinical and Analytical Medicine Göğüs Cerrahisi Hasan Çaylak Göğüs Cerrahisi Metastatik Akciğer Tümörleri Giriş İzole akciğer metastazlarına tedavi edilemez gözüyle bakılmamalıdır Tümör tipine
K TÜMÖRLERDE VE REAKT
 T. C. BAŞKENT ÜNİVERSİTESİ TIP FAKÜLTESİ PATOLOJİ ANABİLİM DALI ASTROSİTİK TÜMÖRLERDE VE REAKTİF GLİOZİS OLGULARINDA CDC25B EKSPRESYONU VE PROGNOSTİK ÖNEMİ: (POTANSİYEL BİR GRADE LEME ARACI OLARAK KULLANILABİLMESİ
T. C. BAŞKENT ÜNİVERSİTESİ TIP FAKÜLTESİ PATOLOJİ ANABİLİM DALI ASTROSİTİK TÜMÖRLERDE VE REAKTİF GLİOZİS OLGULARINDA CDC25B EKSPRESYONU VE PROGNOSTİK ÖNEMİ: (POTANSİYEL BİR GRADE LEME ARACI OLARAK KULLANILABİLMESİ
Beyin Tümörleri: Sınıflama, Evreleme ve Genetik Kavramlar
 Beyin Tümörleri: Sınıflama, Evreleme ve Genetik Kavramlar Prof. Dr. Nail Bulakbaşı Radyoloji & Nöroradyoloji (EDINR, EDiPNR) nail.bulakbasi@kyrenia.edu.tr D R. S U A T G Ü N S E L G İ R N E Ü N İ V E R
Beyin Tümörleri: Sınıflama, Evreleme ve Genetik Kavramlar Prof. Dr. Nail Bulakbaşı Radyoloji & Nöroradyoloji (EDINR, EDiPNR) nail.bulakbasi@kyrenia.edu.tr D R. S U A T G Ü N S E L G İ R N E Ü N İ V E R
MGMT PROMOTER METİLASYONU OLAN GLİOBLASTOMALI OLGULARDA CpG 1, 2, 3 ve 4 METİLASYONUNUN TEDAVİ CEVABINA ETKİSİ
 MGMT PROMOTER METİLASYONU OLAN GLİOBLASTOMALI OLGULARDA CpG 1, 2, 3 ve 4 METİLASYONUNUN TEDAVİ CEVABINA ETKİSİ Dicle Aslan, Oğuz O YıldY ldız, Hilal Akalın, Yagut Akberova, Özlem Canöz, Munis Dündar, D
MGMT PROMOTER METİLASYONU OLAN GLİOBLASTOMALI OLGULARDA CpG 1, 2, 3 ve 4 METİLASYONUNUN TEDAVİ CEVABINA ETKİSİ Dicle Aslan, Oğuz O YıldY ldız, Hilal Akalın, Yagut Akberova, Özlem Canöz, Munis Dündar, D
EĞİTİM - ÖĞRETİM YILI DÖNEM I. III. KURUL DERS PROGRAMI GENETİK BİLGİNİN AKIŞI- DOKUYA GİRİŞ (15 Ocak Mart 2018 )
") 2017 2018 EĞİTİM - ÖĞRETİM YILI DÖNEM I III. KURUL DERS PROGRAMI GENETİK BİLGİNİN AKIŞI- DOKUYA GİRİŞ (15 Ocak 2018-9 Mart 2018 ) Dekan Baş Koordinatör Dönem I Koordinatörü Dönem I Koordinatör Yardımcısı
2017 2018 EĞİTİM - ÖĞRETİM YILI DÖNEM I III. KURUL DERS PROGRAMI GENETİK BİLGİNİN AKIŞI- DOKUYA GİRİŞ (15 Ocak 2018-9 Mart 2018 ) Dekan Baş Koordinatör Dönem I Koordinatörü Dönem I Koordinatör Yardımcısı
Fizyoloji PSİ 123 Hafta Haft 8 a
 Fizyoloji PSİ 123 Hafta 8 Sinir Sisteminin Organizasyonu Sinir Sistemi Merkezi Sinir Sistemi Beyin Omurilik Periferik Sinir Sistemi Merkezi Sinir Sistemi (MSS) Oluşturan Hücreler Ara nöronlar ve motor
Fizyoloji PSİ 123 Hafta 8 Sinir Sisteminin Organizasyonu Sinir Sistemi Merkezi Sinir Sistemi Beyin Omurilik Periferik Sinir Sistemi Merkezi Sinir Sistemi (MSS) Oluşturan Hücreler Ara nöronlar ve motor
YÜKSEK DERECELİ GLİAL TÜMÖR TEDAVİSİNDE GÜNCEL YAKLAŞIM. Dr. Melek Nur Yavuz Akdeniz Üniversitesi Tıp Fakültesi Radyasyon Onkolojisi AD
 YÜKSEK DERECELİ GLİAL TÜMÖR TEDAVİSİNDE GÜNCEL YAKLAŞIM Dr. Melek Nur Yavuz Akdeniz Üniversitesi Tıp Fakültesi Radyasyon Onkolojisi AD Yüksek Dereceli Glial Tümör SSS tümörlerinin %8-12 Anaplastik astrositom
YÜKSEK DERECELİ GLİAL TÜMÖR TEDAVİSİNDE GÜNCEL YAKLAŞIM Dr. Melek Nur Yavuz Akdeniz Üniversitesi Tıp Fakültesi Radyasyon Onkolojisi AD Yüksek Dereceli Glial Tümör SSS tümörlerinin %8-12 Anaplastik astrositom
Göğüs Cerrahisi Alkın Yazıcıoğlu. Journal of Clinical and Analytical Medicine Göğüs Cerrahisi
 Mediastenin Nadir Görülen Tümörleri Tüm mediastinal kitlelerin %10 dan azını meydana getiren bu lezyonlar mezenkimal veya epitelyal kökenli tümörlerden oluşmaktadır. Journal of linical and nalytical Medicine
Mediastenin Nadir Görülen Tümörleri Tüm mediastinal kitlelerin %10 dan azını meydana getiren bu lezyonlar mezenkimal veya epitelyal kökenli tümörlerden oluşmaktadır. Journal of linical and nalytical Medicine
MEME KARSİNOMLARINDA GATA 3 EKSPRESYONU VE KLİNİKOPATOLOJİK PARAMETRELER İLE İLİŞKİSİ
 MEME KARSİNOMLARINDA GATA 3 EKSPRESYONU VE KLİNİKOPATOLOJİK PARAMETRELER İLE İLİŞKİSİ Aslı ÇAKIR 1, Özgür EKİNCİ 2, İpek IŞIK GÖNÜL 2, Bülent ÇETİN 3, Mustafa BENEKLİ 3, Ömer ULUOĞLU 2 1 Çorlu Devlet Hastanesi
MEME KARSİNOMLARINDA GATA 3 EKSPRESYONU VE KLİNİKOPATOLOJİK PARAMETRELER İLE İLİŞKİSİ Aslı ÇAKIR 1, Özgür EKİNCİ 2, İpek IŞIK GÖNÜL 2, Bülent ÇETİN 3, Mustafa BENEKLİ 3, Ömer ULUOĞLU 2 1 Çorlu Devlet Hastanesi
EĞİTİM - ÖĞRETİM YILI DÖNEM I. III. KURULDERS PROGRAMI GENETİK BİLGİNİN AKIŞI - DOKUYA GİRİŞ (15 Ocak Mart 2018 )
") 2017 2018 EĞİTİM - ÖĞRETİM YILI DÖNEM I III. KURULDERS PROGRAMI GENETİK BİLGİNİN AKIŞI - DOKUYA GİRİŞ (15 Ocak 2018-9 Mart 2018 ) Dekan Baş Koordinatör Dönem I Koordinatörü Dönem I Koordinatör Yardımcısı
2017 2018 EĞİTİM - ÖĞRETİM YILI DÖNEM I III. KURULDERS PROGRAMI GENETİK BİLGİNİN AKIŞI - DOKUYA GİRİŞ (15 Ocak 2018-9 Mart 2018 ) Dekan Baş Koordinatör Dönem I Koordinatörü Dönem I Koordinatör Yardımcısı
Beyin tümörü, beyni oluşturan üç bölgeden birinden -beyin, beyincik ve beyin sapıkaynaklanabilir.
 BEYİN TÜMÖRÜ BEYİN TÜMÖRÜ NEDİR? Beyin tümörü beyinde anormal hücrelerin çoğalması sonucu gelişen bir kitledir. Bazı beyin tümörleri iyi huylu (örneğin meningiom), bazıları ise kötü huylu kitlelerdir.
BEYİN TÜMÖRÜ BEYİN TÜMÖRÜ NEDİR? Beyin tümörü beyinde anormal hücrelerin çoğalması sonucu gelişen bir kitledir. Bazı beyin tümörleri iyi huylu (örneğin meningiom), bazıları ise kötü huylu kitlelerdir.
PAPİLLER TİROİD KARSİNOMLU OLGULARIMIZDA BRAF(V600E) GEN MUTASYON ANALİZİ. Klinik ve patolojik özellikler
 GEN MUTASYON ANALİZİ. Klinik ve patolojik özellikler") PAPİLLER TİROİD KARSİNOMLU OLGULARIMIZDA BRAF(V600E) GEN MUTASYON ANALİZİ Klinik ve patolojik özellikler Neslihan KURTULMUŞ,, Mete DÜREN, D Serdar GİRAY, G Ümit İNCE, Önder PEKER, Özlem AYDIN, M.Cengiz
PAPİLLER TİROİD KARSİNOMLU OLGULARIMIZDA BRAF(V600E) GEN MUTASYON ANALİZİ Klinik ve patolojik özellikler Neslihan KURTULMUŞ,, Mete DÜREN, D Serdar GİRAY, G Ümit İNCE, Önder PEKER, Özlem AYDIN, M.Cengiz
Parkinson Hastalığı ile α-sinüklein Geni Polimorfizmlerinin İlişkisinin Araştırılması
 İ.Ü. CERRAHPAŞA TIP FAKÜLTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ ANABİLİM DALI Parkinson Hastalığı ile α-sinüklein Geni Polimorfizmlerinin İlişkisinin Araştırılması Araş.Gör. Yener KURMAN İSTANBUL
İ.Ü. CERRAHPAŞA TIP FAKÜLTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ ANABİLİM DALI Parkinson Hastalığı ile α-sinüklein Geni Polimorfizmlerinin İlişkisinin Araştırılması Araş.Gör. Yener KURMAN İSTANBUL
KOLOREKTAL KANSERLERİN MOLEKÜLER SINIFLAMASI. Doç.Dr.Aytekin AKYOL Hacettepe Üniversitesi Tıp Fakültesi Tıbbi Patoloji Anabilim Dalı 23 Mart 2014
 KOLOREKTAL KANSERLERİN MOLEKÜLER SINIFLAMASI Doç.Dr.Aytekin AKYOL Hacettepe Üniversitesi Tıp Fakültesi Tıbbi Patoloji Anabilim Dalı 23 Mart 2014 Kolorektal Kanserler; Sunum Planı Genel bilgiler Moleküler
KOLOREKTAL KANSERLERİN MOLEKÜLER SINIFLAMASI Doç.Dr.Aytekin AKYOL Hacettepe Üniversitesi Tıp Fakültesi Tıbbi Patoloji Anabilim Dalı 23 Mart 2014 Kolorektal Kanserler; Sunum Planı Genel bilgiler Moleküler
T.C. Sağlık Bakanlığı Şişli Etfal Eğitim ve Araştırma Hastanesi Patoloji Laboratuarı Şef: Doç. Dr. Fevziye Kabukçuoğlu. Dr.
 T.C. Sağlık Bakanlığı Şişli Etfal Eğitim ve Araştırma Hastanesi Patoloji Laboratuarı Şef: Doç. Dr. Fevziye Kabukçuoğlu GRADE IV ASTROSİTİK TÜMÖRLERDE HER2/NEU EKSPRESYONU VE CD105 İLE BOYANAN MİKRODAMAR
T.C. Sağlık Bakanlığı Şişli Etfal Eğitim ve Araştırma Hastanesi Patoloji Laboratuarı Şef: Doç. Dr. Fevziye Kabukçuoğlu GRADE IV ASTROSİTİK TÜMÖRLERDE HER2/NEU EKSPRESYONU VE CD105 İLE BOYANAN MİKRODAMAR
EĞİTİM - ÖĞRETİM YILI DÖNEM I. III. KURULDERS PROGRAMI GENETİK BİLGİNİN AKIŞI- DOKUYA GİRİŞ (16 Ocak Mart 2017 )
") 2015 2016 EĞİTİM - ÖĞRETİM YILI DÖNEM I III. KURULDERS PROGRAMI GENETİK BİLGİNİN AKIŞI- DOKUYA GİRİŞ (16 Ocak 2017-10 Mart 2017 ) Dekan Baş Koordinatör Dönem I Koordinatörü Dönem I Koordinatör Yardımcısı
2015 2016 EĞİTİM - ÖĞRETİM YILI DÖNEM I III. KURULDERS PROGRAMI GENETİK BİLGİNİN AKIŞI- DOKUYA GİRİŞ (16 Ocak 2017-10 Mart 2017 ) Dekan Baş Koordinatör Dönem I Koordinatörü Dönem I Koordinatör Yardımcısı
5. Türk Tıbbi Onkoloji Kongresi
 Bilimsel Program - 20 Mart 2014, Perşembe UĞUR DERMAN SALONU SEÇİLMİŞ VAKA SUNUMLARI - Peritoneal Kanserlerde HIPEC in Yeri HIPEC Nasıl Yapılır? Kolon Kanseri Mezotelyoma KONFERANS - Onkolojide Nereden
Bilimsel Program - 20 Mart 2014, Perşembe UĞUR DERMAN SALONU SEÇİLMİŞ VAKA SUNUMLARI - Peritoneal Kanserlerde HIPEC in Yeri HIPEC Nasıl Yapılır? Kolon Kanseri Mezotelyoma KONFERANS - Onkolojide Nereden
AZ DİFERANSİYE TİROİD KANSERLERİ. Prof. Dr. Müfide Nuran AKÇAY Atatürk Üniversitesi Tıp Fakültesi Genel Cerrahi Anabilim Dalı ERZURUM
 AZ DİFERANSİYE TİROİD KANSERLERİ Prof. Dr. Müfide Nuran AKÇAY Atatürk Üniversitesi Tıp Fakültesi Genel Cerrahi Anabilim Dalı ERZURUM Tanım Az diferansiye tiroid karsinomları, iyi diferansiye ve anaplastik
AZ DİFERANSİYE TİROİD KANSERLERİ Prof. Dr. Müfide Nuran AKÇAY Atatürk Üniversitesi Tıp Fakültesi Genel Cerrahi Anabilim Dalı ERZURUM Tanım Az diferansiye tiroid karsinomları, iyi diferansiye ve anaplastik
HPV Moleküler Tanısında Güncel Durum. DNA bazlı Testler KORAY ERGÜNAY 1.ULUSAL KLİNİK MİKROBİYOLOJİ KONGRESİ
 1.ULUSAL KLİNİK MİKROBİYOLOJİ KONGRESİ HPV Moleküler Tanısında Güncel Durum DNA bazlı Testler KORAY ERGÜNAY Hacettepe Üniversitesi Tıp Fakültesi Tıbbi Mikrobiyoloji AD Viroloji Ünitesi HPV tanısı... Sitolojik/Patolojik
1.ULUSAL KLİNİK MİKROBİYOLOJİ KONGRESİ HPV Moleküler Tanısında Güncel Durum DNA bazlı Testler KORAY ERGÜNAY Hacettepe Üniversitesi Tıp Fakültesi Tıbbi Mikrobiyoloji AD Viroloji Ünitesi HPV tanısı... Sitolojik/Patolojik
Onkolojide Sık Kullanılan Terimler. Yrd.Doç.Dr.Ümmügül Üyetürk 2013
 Onkolojide Sık Kullanılan Terimler Yrd.Doç.Dr.Ümmügül Üyetürk 2013 Kanser Hücrelerin aşırı kontrolsüz üretiminin, bu üretime uygun hücre kaybıyla dengelenemediği, giderek artan hücre kütlelerinin birikimi..
Onkolojide Sık Kullanılan Terimler Yrd.Doç.Dr.Ümmügül Üyetürk 2013 Kanser Hücrelerin aşırı kontrolsüz üretiminin, bu üretime uygun hücre kaybıyla dengelenemediği, giderek artan hücre kütlelerinin birikimi..
Paratiroid Kanserinde Yönetim İzmir den Üç Merkezli Deneyim
 Paratiroid Kanserinde Yönetim İzmir den Üç Merkezli Deneyim Murat Özdemir, Özer Makay, Tevfik Demir*, Berk Göktepe, Kamil Erözkan, Barış Bingül**, Yeşim Ertan**, Hüsnü Buğdaycı***, Gökhan İçöz, Mahir Akyıldız
Paratiroid Kanserinde Yönetim İzmir den Üç Merkezli Deneyim Murat Özdemir, Özer Makay, Tevfik Demir*, Berk Göktepe, Kamil Erözkan, Barış Bingül**, Yeşim Ertan**, Hüsnü Buğdaycı***, Gökhan İçöz, Mahir Akyıldız
SİNİR SİSTEMİ Sinir sistemi vücutta, kas kontraksiyonlarını, hızlı değişen viseral olayları ve bazı endokrin bezlerin sekresyon hızlarını kontrol eder
 SİNİR SİSTEMİ SİNİR SİSTEMİ Sinir sistemi vücutta, kas kontraksiyonlarını, hızlı değişen viseral olayları ve bazı endokrin bezlerin sekresyon hızlarını kontrol eder. Çeşitli duyu organlarından milyonlarca
SİNİR SİSTEMİ SİNİR SİSTEMİ Sinir sistemi vücutta, kas kontraksiyonlarını, hızlı değişen viseral olayları ve bazı endokrin bezlerin sekresyon hızlarını kontrol eder. Çeşitli duyu organlarından milyonlarca
Savaş Baba, Sabri Özden, Barış Saylam, Umut Fırat Turan Ankara Numune EAH. Meme Endokrin Cerrahi Kliniği
 Savaş Baba, Sabri Özden, Barış Saylam, Umut Fırat Turan Ankara Numune EAH. Meme Endokrin Cerrahi Kliniği Tiroid kanserleri bütün kanserler içinde yaklaşık % 1 oranında görülmekte olup, özellikle kadınlarda
Savaş Baba, Sabri Özden, Barış Saylam, Umut Fırat Turan Ankara Numune EAH. Meme Endokrin Cerrahi Kliniği Tiroid kanserleri bütün kanserler içinde yaklaşık % 1 oranında görülmekte olup, özellikle kadınlarda
BCC DE GÜNCEL Prof. Dr. Kamer GÜNDÜZ
 BCC DE GÜNCEL Prof. Dr. Kamer GÜNDÜZ Celal Bayar Üniversitesi Deri ve Zührevi Hastalıklar Anabilim Dalı-MANİSA Bazal Hücreli Kanser (BCC) 1827 - Arthur Jacob En sık rastlanan deri kanseri (%70-80) Açık
BCC DE GÜNCEL Prof. Dr. Kamer GÜNDÜZ Celal Bayar Üniversitesi Deri ve Zührevi Hastalıklar Anabilim Dalı-MANİSA Bazal Hücreli Kanser (BCC) 1827 - Arthur Jacob En sık rastlanan deri kanseri (%70-80) Açık
1. Oturum: Meme Kanserine Giriş, Patoloji ve Alt Tiplendirme Oturum Başkanları : Dr. Orhan ŞENCAN, Dr. İrfan ÇİÇİN
 TTOD Meme Kanseri Güncelleme Kursu 13-14 Haziran 2015 İstanbul 08:25-08:30 Açılış Dr. Cem BÖRÜBAN Oturum Başkanları : Dr. Fikri İÇLİ, Dr. Gül Başaran, Dr. Erhan GÖKMEN 08:00-08:30 Pratiği Değiştiren Çalışmalar.
TTOD Meme Kanseri Güncelleme Kursu 13-14 Haziran 2015 İstanbul 08:25-08:30 Açılış Dr. Cem BÖRÜBAN Oturum Başkanları : Dr. Fikri İÇLİ, Dr. Gül Başaran, Dr. Erhan GÖKMEN 08:00-08:30 Pratiği Değiştiren Çalışmalar.
Adölesanda Lösemi & İnfant Lösemi
 Adölesanda Lösemi & İnfant Lösemi Prof. Dr. Özcan Bör Eskişehir Osmangazi Üniversitesi TPHD OKULU 18 20 Kasım 2016 Ankara 1 Adölesanda Lösemi Dünya Sağlık Örgütü 10 19 yaşlarını Adölesan Dönemi olarak
Adölesanda Lösemi & İnfant Lösemi Prof. Dr. Özcan Bör Eskişehir Osmangazi Üniversitesi TPHD OKULU 18 20 Kasım 2016 Ankara 1 Adölesanda Lösemi Dünya Sağlık Örgütü 10 19 yaşlarını Adölesan Dönemi olarak
Dr. Yaşar BEDÜK Ankara Üniversitesi Tıp Fakültesi
 Dr. Yaşar BEDÜK Ankara Üniversitesi Tıp Fakültesi Testis tümörleri Erkeklerdeki kanserlerin %1 i Yıllık %3-6 artış Avrupa da her yıl 24.000 yeni vak a Testis Tümör Epidemiyolojisi İnsidans dünya genelinde
Dr. Yaşar BEDÜK Ankara Üniversitesi Tıp Fakültesi Testis tümörleri Erkeklerdeki kanserlerin %1 i Yıllık %3-6 artış Avrupa da her yıl 24.000 yeni vak a Testis Tümör Epidemiyolojisi İnsidans dünya genelinde
MEME PATOLOJİSİ Slayt Semineri Oturum Başkanları: Dr. Osman ZEKİOĞLU - Dr. Gülnur GÜLER
 MEME PATOLOJİSİ Slayt Semineri Oturum Başkanları: Dr. Osman ZEKİOĞLU - Dr. Gülnur GÜLER OLGU SUNUMU Dr Tülin Öztürk İ.Ü. Cerrahpaşa Tıp Fakültesi Patoloji Anabilim Dalı 25. Ulusal Patoloji Kongresi 14-17
MEME PATOLOJİSİ Slayt Semineri Oturum Başkanları: Dr. Osman ZEKİOĞLU - Dr. Gülnur GÜLER OLGU SUNUMU Dr Tülin Öztürk İ.Ü. Cerrahpaşa Tıp Fakültesi Patoloji Anabilim Dalı 25. Ulusal Patoloji Kongresi 14-17
KOLOREKTAL KARSİNOMLU HASTALARDA PRİMER İLE METASTAZ ARASINDA KRAS DİSKORDANSI
 KOLOREKTAL KARSİNOMLU HASTALARDA PRİMER İLE METASTAZ ARASINDA KRAS DİSKORDANSI AHMET ŞİYAR EKİNCİ1, UMUT DEMİRCİ 1, BERNA ÖKSÜZOĞLU1, AYŞEGÜL ÖZTÜRK2, ONUR EŞBAH1, TAHSİN ÖZATLI1, ÖZNUR BAL1, AYŞE DEMİRCİ1,
KOLOREKTAL KARSİNOMLU HASTALARDA PRİMER İLE METASTAZ ARASINDA KRAS DİSKORDANSI AHMET ŞİYAR EKİNCİ1, UMUT DEMİRCİ 1, BERNA ÖKSÜZOĞLU1, AYŞEGÜL ÖZTÜRK2, ONUR EŞBAH1, TAHSİN ÖZATLI1, ÖZNUR BAL1, AYŞE DEMİRCİ1,
AKCİĞERİN NÖROENDOKRİN TÜMÖRLERİ. Doç. Dr. Mutlu DEMİRAY Bursa Medical Park Hastanesi
 AKCİĞERİN NÖROENDOKRİN TÜMÖRLERİ Doç. Dr. Mutlu DEMİRAY Bursa Medical Park Hastanesi Nöroendokrin tümörlerde 2004 WHO sınıflaması Tümör Tipi Tipik Karsinoid Atipik Karsinoid Büyük Hücreli nöroendokrin
AKCİĞERİN NÖROENDOKRİN TÜMÖRLERİ Doç. Dr. Mutlu DEMİRAY Bursa Medical Park Hastanesi Nöroendokrin tümörlerde 2004 WHO sınıflaması Tümör Tipi Tipik Karsinoid Atipik Karsinoid Büyük Hücreli nöroendokrin
TAKD olgu sunumları- 21 Kasım Dr Şebnem Batur Dr Büge ÖZ İÜ Cerrahpaşa Tıp Fakültesi Patoloji AD
 TAKD olgu sunumları- 21 Kasım 2012 Dr Şebnem Batur Dr Büge ÖZ İÜ Cerrahpaşa Tıp Fakültesi Patoloji AD Konuşma akışı; ALK mutasyonu değerlendirmedeki sorunlar ROS-1 mutasyonu Avrupa pulmoner patoloji çalışma
TAKD olgu sunumları- 21 Kasım 2012 Dr Şebnem Batur Dr Büge ÖZ İÜ Cerrahpaşa Tıp Fakültesi Patoloji AD Konuşma akışı; ALK mutasyonu değerlendirmedeki sorunlar ROS-1 mutasyonu Avrupa pulmoner patoloji çalışma
T.C. ÇUKUROVA ÜNİVERSİTESİ TIP FAKÜLTESİ TIBBİ PATOLOJİ ANABİLİM DALI
 T.C. ÇUKUROVA ÜNİVERSİTESİ TIP FAKÜLTESİ TIBBİ PATOLOJİ ANABİLİM DALI GLİOBLASTOMLARDA METİL GUANİN METİL TRANSFERAZ PROMOTER BÖLGESİNDE METİLASYONUN POLİMERAZ ZİNCİR REAKSİYON YÖNTEMİYLE GÖSTERİLMESİ,
T.C. ÇUKUROVA ÜNİVERSİTESİ TIP FAKÜLTESİ TIBBİ PATOLOJİ ANABİLİM DALI GLİOBLASTOMLARDA METİL GUANİN METİL TRANSFERAZ PROMOTER BÖLGESİNDE METİLASYONUN POLİMERAZ ZİNCİR REAKSİYON YÖNTEMİYLE GÖSTERİLMESİ,
Cebinizdeki Görüntü Analizi. Ki67 Proliferasyon İndeksini Belirlemek İçin Pratik Ve Basit Uygulama. Dr. Haldun Umudum Ufuk Üniversitesi, Ankara
 Cebinizdeki Görüntü Analizi. Ki67 Proliferasyon İndeksini Belirlemek İçin Pratik Ve Basit Uygulama. Dr. Haldun Umudum Ufuk Üniversitesi, Ankara Giriş: Günlük patoloji pratiğinde Ki67 pozitif hücrelerinin
Cebinizdeki Görüntü Analizi. Ki67 Proliferasyon İndeksini Belirlemek İçin Pratik Ve Basit Uygulama. Dr. Haldun Umudum Ufuk Üniversitesi, Ankara Giriş: Günlük patoloji pratiğinde Ki67 pozitif hücrelerinin
Küçük Hücreli Dışı Akciğer Karsinomlarının EGFR Mutasyon Analizinde Real-Time PCR Yöntemi ile Mutasyona Spesifik İmmünohistokimyanın Karşılaştırılması
 Küçük Hücreli Dışı Akciğer Karsinomlarının EGFR Mutasyon Analizinde Real-Time PCR Yöntemi ile Mutasyona Spesifik nın Karşılaştırılması Dr.M.Çisel Aydın, Doç.Dr.Sevgen Önder, Prof.Dr.Gaye Güler Tezel Hacettepe
Küçük Hücreli Dışı Akciğer Karsinomlarının EGFR Mutasyon Analizinde Real-Time PCR Yöntemi ile Mutasyona Spesifik nın Karşılaştırılması Dr.M.Çisel Aydın, Doç.Dr.Sevgen Önder, Prof.Dr.Gaye Güler Tezel Hacettepe
İkincil Kanserler SUNA EMİR. Ankara Çocuk Sağlığı ve Hastalıkları Hematoloji Onkoloji EAH Çocuk Hematoloji Onkoloji Kliniği
 İkincil Kanserler SUNA EMİR Ankara Çocuk Sağlığı ve Hastalıkları Hematoloji Onkoloji EAH Çocuk Hematoloji Onkoloji Kliniği Sunum akışı İkincil kanser tanımı Neden önemli bir konu? İkincil kanser oluşumunda
İkincil Kanserler SUNA EMİR Ankara Çocuk Sağlığı ve Hastalıkları Hematoloji Onkoloji EAH Çocuk Hematoloji Onkoloji Kliniği Sunum akışı İkincil kanser tanımı Neden önemli bir konu? İkincil kanser oluşumunda
Genel Kimya II (2 0 4)
") Genel Kimya II (2 0 4) 1 Kimyasal Kinetik 2 Kimyasal Dengenin İlkeleri I 3 Kimyasal Dengenin İlkeleri II 4 Asitler ve Bazlar I 5 Asitler Bazlar II 6 Sulu Çözelti Reaksiyonları ve İyon Dengeleri I 7 Sulu
Genel Kimya II (2 0 4) 1 Kimyasal Kinetik 2 Kimyasal Dengenin İlkeleri I 3 Kimyasal Dengenin İlkeleri II 4 Asitler ve Bazlar I 5 Asitler Bazlar II 6 Sulu Çözelti Reaksiyonları ve İyon Dengeleri I 7 Sulu
b. Amaç: Gen anatomisi ile ilgili genel bilgi öğretilmesi amaçlanmıştır.
 TIBBİ GENETİK I-DERS TANIMLARI 1-Tanım: DNA ve RNA yapısının öğretilmesi. b. Amaç: DNA nın genetik materyal olmasında moleküler yapısının önemi ve RNA yapısının proteine geçiş ve gen ekspresyonu kontrolündeki
TIBBİ GENETİK I-DERS TANIMLARI 1-Tanım: DNA ve RNA yapısının öğretilmesi. b. Amaç: DNA nın genetik materyal olmasında moleküler yapısının önemi ve RNA yapısının proteine geçiş ve gen ekspresyonu kontrolündeki
Epidermal bazal hücrelerden veya kıl folikülünün dış kök kılıfından köken alan malin deri tm
 BAZAL HÜCRELİ KARSİNOM Epidermal bazal hücrelerden veya kıl folikülünün dış kök kılıfından köken alan malin deri tm Nadiren met. yapar fakat tedavisiz bırakıldığında invazif davranış göstermesi,lokal invazyon,
BAZAL HÜCRELİ KARSİNOM Epidermal bazal hücrelerden veya kıl folikülünün dış kök kılıfından köken alan malin deri tm Nadiren met. yapar fakat tedavisiz bırakıldığında invazif davranış göstermesi,lokal invazyon,
MEME PATOLOJİSİ SLAYT SEMİNERİ
 MEME PATOLOJİSİ SLAYT SEMİNERİ Prof. Dr. Şahsine Tolunay Uludağ Üniversitesi Tıp Fakültesi Patoloji Anabilim Dalı 17.10.2015 OLGU 43 yaşında kadın 2 çocuğu var Sol memede ağrı ve kitle yakınması mevcut
MEME PATOLOJİSİ SLAYT SEMİNERİ Prof. Dr. Şahsine Tolunay Uludağ Üniversitesi Tıp Fakültesi Patoloji Anabilim Dalı 17.10.2015 OLGU 43 yaşında kadın 2 çocuğu var Sol memede ağrı ve kitle yakınması mevcut
(ZORUNLU) MOLEKÜLER İMMÜNOLOJİ I (TBG 607 TEORİK 3, 3 KREDİ)
 MOLEKÜLER İMMÜNOLOJİ I (TBG 607 TEORİK 3, 3 KREDİ)") T. C. İSTANBUL BİLİM ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ VE GENETİK ANABİLİM DALI YÜKSEK LİSANS PROGRAMI 2015-2016 EĞİTİM-ÖĞRETİM YILI DERS İÇERİKLERİ I. YARIYIL (ZORUNLU) MOLEKÜLER
T. C. İSTANBUL BİLİM ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ VE GENETİK ANABİLİM DALI YÜKSEK LİSANS PROGRAMI 2015-2016 EĞİTİM-ÖĞRETİM YILI DERS İÇERİKLERİ I. YARIYIL (ZORUNLU) MOLEKÜLER
Tıbbın Geleceğine dair.. Genetik Testler ve Kişiselleşmiş Tıp Anlayışı. B. Aysin Sermen
 Tıbbın Geleceğine dair.. Genetik Testler ve Kişiselleşmiş Tıp Anlayışı B. Aysin Sermen Daha güçlü.. Daha atletik.. Daha genç.. Daha huzurlu.. Daha mutlu.. Daha akıllı.. Daha sağlıklı.. Daha akıllı ve sağlıklı
Tıbbın Geleceğine dair.. Genetik Testler ve Kişiselleşmiş Tıp Anlayışı B. Aysin Sermen Daha güçlü.. Daha atletik.. Daha genç.. Daha huzurlu.. Daha mutlu.. Daha akıllı.. Daha sağlıklı.. Daha akıllı ve sağlıklı
Türk Nöroşirurji Akademisi
 Türk Nöroşirürji Akademisi nin Saygıdeğer Üyeleri, TNA Yönetim Kurulu Prof. Dr. M Necmettin Pamir, Başkan Prof. Dr. Tunçalp Özgen, Önceki Başkan Prof. Dr. Kaya Aksoy, Gelecek Dönem Başkanı Prof. Dr. Uygur
Türk Nöroşirürji Akademisi nin Saygıdeğer Üyeleri, TNA Yönetim Kurulu Prof. Dr. M Necmettin Pamir, Başkan Prof. Dr. Tunçalp Özgen, Önceki Başkan Prof. Dr. Kaya Aksoy, Gelecek Dönem Başkanı Prof. Dr. Uygur
Konuşma planı PEDİATRİK SUPRATENTORYAL BEYİN TÜMÖRLERİ. Epidemiyoloji. CBTRUS 2011 report. Genetik sendromlar 4/26/2014
 Konuşma planı PEDİATRİK SUPRATENTORYAL BEYİN TÜMÖRLERİ Dr. Simay Altan Kara Acıbadem Üniversitesi Tıp Fakültesi Radyoloji AD Epidemiyoloji Klinik Konvansiyonel görüntüleme İleri MR teknikleri Tümörler
Konuşma planı PEDİATRİK SUPRATENTORYAL BEYİN TÜMÖRLERİ Dr. Simay Altan Kara Acıbadem Üniversitesi Tıp Fakültesi Radyoloji AD Epidemiyoloji Klinik Konvansiyonel görüntüleme İleri MR teknikleri Tümörler
HALK SAĞLIĞI ANABĠLĠM DALI. Ders adı : Endokrin çevre bozucular ve tarama programı
 HALK SAĞLIĞI ANABĠLĠM DALI Ders adı : Endokrin çevre bozucular ve tarama programı Öğretim Üyesi : Prof. Dr. A. Emel ÖNAL Endokrin sistemin çalışmasını değiştiren, sağlıklı insanda veya çocuklarında sağlık
HALK SAĞLIĞI ANABĠLĠM DALI Ders adı : Endokrin çevre bozucular ve tarama programı Öğretim Üyesi : Prof. Dr. A. Emel ÖNAL Endokrin sistemin çalışmasını değiştiren, sağlıklı insanda veya çocuklarında sağlık
Replikasyon, Transkripsiyon ve Translasyon. Yrd. Doç. Dr. Osman İBİŞ
 Replikasyon, Transkripsiyon ve Translasyon Yrd. Doç. Dr. Osman İBİŞ DNA replikasyonu DNA nın replikasyonu, DNA molekülünün, sakladığı genetik bilgilerin sonraki nesillere aktarılması için kendi kopyasını
Replikasyon, Transkripsiyon ve Translasyon Yrd. Doç. Dr. Osman İBİŞ DNA replikasyonu DNA nın replikasyonu, DNA molekülünün, sakladığı genetik bilgilerin sonraki nesillere aktarılması için kendi kopyasını
Dr. A. YÜKSEL BARUT 1
 1 1 Santral Sinir sistemi (SSS): Beyin (Encephalon) ve Omurilik (Medulla Spinalis), 2 Periferik Sinir Sistemi (PSS) : Beyin sapı ve omurilikten çıkan bütün sinirler, 3 Otonom sinir sistemi (OSS) : Hem
1 1 Santral Sinir sistemi (SSS): Beyin (Encephalon) ve Omurilik (Medulla Spinalis), 2 Periferik Sinir Sistemi (PSS) : Beyin sapı ve omurilikten çıkan bütün sinirler, 3 Otonom sinir sistemi (OSS) : Hem
T. C. MUĞLA SITKI KOÇMAN ÜNİVERSİTESİ TIP FAKÜLTESİ 2014 2015 EĞİTİM - ÖĞRETİM YILI DÖNEM III
 T. C. MUĞLA SITKI KOÇMAN ÜNİVERSİTESİ TIP FAKÜLTESİ 2014 2015 EĞİTİM - ÖĞRETİM YILI DÖNEM III DOKU ZEDELENMESI VE ENFEKSIYON DERS KURULU ( 1. ) DERS KURULU (15 EYLÜL- 24 EKİM 2014) DERS PROGRAMI T. C.
T. C. MUĞLA SITKI KOÇMAN ÜNİVERSİTESİ TIP FAKÜLTESİ 2014 2015 EĞİTİM - ÖĞRETİM YILI DÖNEM III DOKU ZEDELENMESI VE ENFEKSIYON DERS KURULU ( 1. ) DERS KURULU (15 EYLÜL- 24 EKİM 2014) DERS PROGRAMI T. C.
TURNOG Yeni olan ne var sempozyumu
 Nöroonkolojik Cerrahi Öğretim ve Eğitim Grubu TURNOG Yeni olan ne var sempozyumu 23-24 Aralık 2011 Barbaros Point Hotel, Gayrettepe, İstanbul YÖNETİM KURULU MESAJI Türk Nöroşirürji Derneği Yönetim Kurulu
Nöroonkolojik Cerrahi Öğretim ve Eğitim Grubu TURNOG Yeni olan ne var sempozyumu 23-24 Aralık 2011 Barbaros Point Hotel, Gayrettepe, İstanbul YÖNETİM KURULU MESAJI Türk Nöroşirürji Derneği Yönetim Kurulu
ERİŞKİN BEYİN TÜMÖRLERİ. Yrd.Doç.Dr.Adalet ARIKANOĞLU D.Ü.T.F.Nöroloji A.B.D
 ERİŞKİN BEYİN TÜMÖRLERİ Yrd.Doç.Dr.Adalet ARIKANOĞLU D.Ü.T.F.Nöroloji A.B.D İnsidansı Primer beyin tümörleri 6/100.000/yıl görülür PBT hastaların % 10 u çocukluk çağında görülür. Yerleşim yeri ve histolojik
ERİŞKİN BEYİN TÜMÖRLERİ Yrd.Doç.Dr.Adalet ARIKANOĞLU D.Ü.T.F.Nöroloji A.B.D İnsidansı Primer beyin tümörleri 6/100.000/yıl görülür PBT hastaların % 10 u çocukluk çağında görülür. Yerleşim yeri ve histolojik
Prostat Tümörlerinde WHO 2016 Sınıflandırması DR. BORA GÜREL KOCAELİ ÜNİVERSİTESİ TIP FAKÜLTESİ PATOLOJİ ANABİLİM DALI
 Prostat Tümörlerinde WHO 2016 Sınıflandırması DR. BORA GÜREL KOCAELİ ÜNİVERSİTESİ TIP FAKÜLTESİ PATOLOJİ ANABİLİM DALI Giriş En son WHO sınıflandırması 2004 yılında Son 12 yılda hatırı sayılır yenilikler
Prostat Tümörlerinde WHO 2016 Sınıflandırması DR. BORA GÜREL KOCAELİ ÜNİVERSİTESİ TIP FAKÜLTESİ PATOLOJİ ANABİLİM DALI Giriş En son WHO sınıflandırması 2004 yılında Son 12 yılda hatırı sayılır yenilikler
Mikrobiyolojide Moleküler Tanı Yöntemleri. Dr.Tuncer ÖZEKİNCİ Dicle Üniversitesi Tıp Fakültesi Tıbbi Mikrobiyoloji A.D
 Mikrobiyolojide Moleküler Tanı Yöntemleri Dr.Tuncer ÖZEKİNCİ Dicle Üniversitesi Tıp Fakültesi Tıbbi Mikrobiyoloji A.D 1 Enfeksiyonun Özgül Laboratuvar Tanısı Mikroorganizmanın üretilmesi Mikroorganizmaya
Mikrobiyolojide Moleküler Tanı Yöntemleri Dr.Tuncer ÖZEKİNCİ Dicle Üniversitesi Tıp Fakültesi Tıbbi Mikrobiyoloji A.D 1 Enfeksiyonun Özgül Laboratuvar Tanısı Mikroorganizmanın üretilmesi Mikroorganizmaya
BİRİNCİL KEMİK KANSERİ
 BİRİNCİL KEMİK KANSERİ KONDROSARKOM (KS) PROF. DR. LEVENT ERALP Ortopedi ve Travmatoloji Uzmanı İÇİNDEKİLER Kondrosarkom Nedir? KS dan kimler etkilenir? Bulgular nelerdir? KS tipleri nelerdir? Risk faktörleri
BİRİNCİL KEMİK KANSERİ KONDROSARKOM (KS) PROF. DR. LEVENT ERALP Ortopedi ve Travmatoloji Uzmanı İÇİNDEKİLER Kondrosarkom Nedir? KS dan kimler etkilenir? Bulgular nelerdir? KS tipleri nelerdir? Risk faktörleri
Meme ve Over Kanserlerinde Laboratuvar: Klinisyenin Laboratuvardan Beklentisi
 Meme ve Over Kanserlerinde Laboratuvar: Klinisyenin Laboratuvardan Beklentisi Dr. Handan Onur XXI. Düzen Klinik Laboratuvar Günleri, Ankara, 23 Ekim 2011 MEME KANSERİ Meme Kanseri Sıklıkla meme başına
Meme ve Over Kanserlerinde Laboratuvar: Klinisyenin Laboratuvardan Beklentisi Dr. Handan Onur XXI. Düzen Klinik Laboratuvar Günleri, Ankara, 23 Ekim 2011 MEME KANSERİ Meme Kanseri Sıklıkla meme başına
TIBBĠ BĠLĠMLERE GĠRĠġ DĠLĠMĠ MĠKROBĠYOLOJĠ ANABĠLĠM DALI
 TIBBĠ BĠLĠMLERE GĠRĠġ DĠLĠMĠ MĠKROBĠYOLOJĠ ANABĠLĠM DALI ÖĞRETĠM ÜYESĠ : Prof. Dr. O. ġadi Yenen Ders: VĠROLOJĠYE GĠRĠġ, TARĠHÇE ve EVRĠM 1. Virusların tanımlanması ve rolüne ilişkin önemli tarihsel gelişmelerin
TIBBĠ BĠLĠMLERE GĠRĠġ DĠLĠMĠ MĠKROBĠYOLOJĠ ANABĠLĠM DALI ÖĞRETĠM ÜYESĠ : Prof. Dr. O. ġadi Yenen Ders: VĠROLOJĠYE GĠRĠġ, TARĠHÇE ve EVRĠM 1. Virusların tanımlanması ve rolüne ilişkin önemli tarihsel gelişmelerin
Dr. Nilay HAKAN Muğla Sıtkı Koçman Üniv. Tıp Fak. Neonatoloji Bilim Dalı Perinatal Medicine Nisan 2017, İZMİR
 Nedeni Açıklanamayan Patolojik ve Uzamış Sarılığı Olan Yenidoğanlarda UDP- Glukuroniltransferaz 1 (UGT1A1), Hepatik Organik Anyon Taşıyıcısı (SLCO1B) ve Glutatyon S-transferaz (GST) Polimorfizmlerinin
Nedeni Açıklanamayan Patolojik ve Uzamış Sarılığı Olan Yenidoğanlarda UDP- Glukuroniltransferaz 1 (UGT1A1), Hepatik Organik Anyon Taşıyıcısı (SLCO1B) ve Glutatyon S-transferaz (GST) Polimorfizmlerinin
Rastgele (Stokas7k) kanser modeli - Tümör içindeki her hücre yeni bir kanseri başla5r
 kanser modeli - Tümör içindeki her hücre yeni bir kanseri başla5r") Kanser Kök Hücre Kanser Modelleri Rastgele (Stokas7k) kanser modeli - Tümör içindeki her hücre yeni bir kanseri başla5r Kök hücre (Hiyerarşi) modeli - Tümör içindeki bazı hücreler yeni bir kanseri başla5r
Kanser Kök Hücre Kanser Modelleri Rastgele (Stokas7k) kanser modeli - Tümör içindeki her hücre yeni bir kanseri başla5r Kök hücre (Hiyerarşi) modeli - Tümör içindeki bazı hücreler yeni bir kanseri başla5r
Malignite ve Transplantasyon. Doç. Dr. Halil Yazıcı İstanbul Tıp Fakültesi Nefroloji Bilim Dalı
 Malignite ve Transplantasyon Doç. Dr. Halil Yazıcı İstanbul Tıp Fakültesi Nefroloji Bilim Dalı Sunum Planı -Pretransplant malignitesi olan alıcı -Pretransplant malignitesi olan donör -Posttransplant de
Malignite ve Transplantasyon Doç. Dr. Halil Yazıcı İstanbul Tıp Fakültesi Nefroloji Bilim Dalı Sunum Planı -Pretransplant malignitesi olan alıcı -Pretransplant malignitesi olan donör -Posttransplant de
KANSER NEDİR? ONKOGEN VE KANSER. Hücre döngüsü. Siklin-Siklin Kinaz 1/30/2012 HÜCRE DÖNGÜSÜ. Siklin Kinaz inhibitörleri BÜYÜME FAKTÖRLERİ
 KANSER NEDİR? ONKOGEN VE KANSER Prof.Dr.Dildar Konukoğlu Bir hücre veya hücre grubunun kontrol dışı büyümesi ve çoğalması ve Bu hücrelerin bulundukları yerden ayrılarak farklı lokalizasyonlarda bu faaliyetlerini
KANSER NEDİR? ONKOGEN VE KANSER Prof.Dr.Dildar Konukoğlu Bir hücre veya hücre grubunun kontrol dışı büyümesi ve çoğalması ve Bu hücrelerin bulundukları yerden ayrılarak farklı lokalizasyonlarda bu faaliyetlerini
EĞİTİM - ÖĞRETİM YILI DÖNEM II I. KURUL DERS PROGRAMI (SĠNĠR SĠSTEMĠ) 18 EYLÜL KASIM 2017 (8 HAFTA)
 18 EYLÜL KASIM 2017 (8 HAFTA)") 2017-2018 EĞİTİM - ÖĞRETİM YILI DÖNEM II I. KURUL DERS PROGRAMI (SĠNĠR SĠSTEMĠ) 18 EYLÜL 2017 10 KASIM 2017 (8 HAFTA) DEKAN BAŞKOORDİNATÖR DÖNEM II KOORDİNATÖRÜ DÖNEM II KOORDİNATÖR YARDIMCISI DÖNEM II
2017-2018 EĞİTİM - ÖĞRETİM YILI DÖNEM II I. KURUL DERS PROGRAMI (SĠNĠR SĠSTEMĠ) 18 EYLÜL 2017 10 KASIM 2017 (8 HAFTA) DEKAN BAŞKOORDİNATÖR DÖNEM II KOORDİNATÖRÜ DÖNEM II KOORDİNATÖR YARDIMCISI DÖNEM II
KANSERDE RADYOLOJİK GÖRÜNTÜLEME DOÇ. DR.İSMAİL MİHMANLI
 KANSERDE RADYOLOJİK GÖRÜNTÜLEME DOÇ. DR.İSMAİL MİHMANLI AMAÇ Kanser ön ya da kesin tanılı hastalarda radyolojik algoritmayı belirlemek ÖĞRENİM HEDEFLERİ Kanser riski olan hastalara doğru radyolojik tetkik
KANSERDE RADYOLOJİK GÖRÜNTÜLEME DOÇ. DR.İSMAİL MİHMANLI AMAÇ Kanser ön ya da kesin tanılı hastalarda radyolojik algoritmayı belirlemek ÖĞRENİM HEDEFLERİ Kanser riski olan hastalara doğru radyolojik tetkik
OLİGODENDROGLİOM VE OLİGOASTROSİTOMLARDA GENETİK RİSK FAKTÖRLERİ VE BU FAKTÖRLERİN PROGNOZ ÜZERİNE OLAN ETKİLERİ
 T.C ÇUKUROVA ÜNİVERSİTESİ TIP FAKÜLTESİ BEYİN VE SİNİR CERRAHİSİ ANABİLİM DALI OLİGODENDROGLİOM VE OLİGOASTROSİTOMLARDA GENETİK RİSK FAKTÖRLERİ VE BU FAKTÖRLERİN PROGNOZ ÜZERİNE OLAN ETKİLERİ Dr. MEHMET
T.C ÇUKUROVA ÜNİVERSİTESİ TIP FAKÜLTESİ BEYİN VE SİNİR CERRAHİSİ ANABİLİM DALI OLİGODENDROGLİOM VE OLİGOASTROSİTOMLARDA GENETİK RİSK FAKTÖRLERİ VE BU FAKTÖRLERİN PROGNOZ ÜZERİNE OLAN ETKİLERİ Dr. MEHMET
İyonize Radyasyonun Hücresel Düzeydeki Etkileri ve Moleküler Yaklaşımlar
 İyonize Radyasyonun Hücresel Düzeydeki Etkileri ve Moleküler Yaklaşımlar Aysun Manisalıgil, Ayşegül Yurt Dokuz Eylül Üniversitesi Sağlık Bilimleri Enstitüsü Medikal Fizik Anabilim Dalı Hücre ve Moleküller
İyonize Radyasyonun Hücresel Düzeydeki Etkileri ve Moleküler Yaklaşımlar Aysun Manisalıgil, Ayşegül Yurt Dokuz Eylül Üniversitesi Sağlık Bilimleri Enstitüsü Medikal Fizik Anabilim Dalı Hücre ve Moleküller
Over Kanserinde Tedavi. Dr. M. Faruk Köse Etlik Zübeyde Hanım Kadın Hastalıkları Eğitim ve Araştırma Hastanesi
 Over Kanserinde Tedavi Dr. M. Faruk Köse Etlik Zübeyde Hanım Kadın Hastalıkları Eğitim ve Araştırma Hastanesi Over Ca Tipleri Tip 1 Tip 2 Yavaş ilerleyen İyi belirlenmiş borderline prekürsör lezyonları
Over Kanserinde Tedavi Dr. M. Faruk Köse Etlik Zübeyde Hanım Kadın Hastalıkları Eğitim ve Araştırma Hastanesi Over Ca Tipleri Tip 1 Tip 2 Yavaş ilerleyen İyi belirlenmiş borderline prekürsör lezyonları
MOLEKÜLER TANISI DÜZEN GENETİK HASTALIKLAR TANI MERKEZİ. SERPİL ERASLAN, PhD
 β-talaseminin MOLEKÜLER TANISI DÜZEN GENETİK HASTALIKLAR TANI MERKEZİ SERPİL ERASLAN, PhD BETA TALASEMİ HEMOGLOBİNOPATİLER Otozomal resesif (globin gen ailesi) Özellikle Çukurova, Akdeniz kıyı şeridi,
β-talaseminin MOLEKÜLER TANISI DÜZEN GENETİK HASTALIKLAR TANI MERKEZİ SERPİL ERASLAN, PhD BETA TALASEMİ HEMOGLOBİNOPATİLER Otozomal resesif (globin gen ailesi) Özellikle Çukurova, Akdeniz kıyı şeridi,
Pleomorfik mezankimal tümörler ve taklitçileri. Dr. Bahar Müezzinoğlu
 Pleomorfik mezankimal tümörler ve taklitçileri Dr. Bahar Müezzinoğlu Pleomorfik lezyon yaş yerleşim hikaye Pleomorfik lezyon neoplastik Neoplastik değil benign malign sarkom Karsinom Lenfoma melanom Pleomorfizm
Pleomorfik mezankimal tümörler ve taklitçileri Dr. Bahar Müezzinoğlu Pleomorfik lezyon yaş yerleşim hikaye Pleomorfik lezyon neoplastik Neoplastik değil benign malign sarkom Karsinom Lenfoma melanom Pleomorfizm
Santral (merkezi) sinir sistemi
 sinir sistemi") Santral (merkezi) sinir sistemi 1 2 Beyin birçok dokunun kontrollerini üstlenmiştir. Çalışması hakkında hala yeterli veri edinemediğimiz beyin, hafıza ve karar verme organı olarak kabul edilir. Sadece
Santral (merkezi) sinir sistemi 1 2 Beyin birçok dokunun kontrollerini üstlenmiştir. Çalışması hakkında hala yeterli veri edinemediğimiz beyin, hafıza ve karar verme organı olarak kabul edilir. Sadece
TİROİD NODÜLLERİNİN SIVI BAZLI (SurePath) SİTOLOJİSİ: 3 Yıllık Histopatolojik Korelasyon
 SİTOLOJİSİ: 3 Yıllık Histopatolojik Korelasyon") TİROİD NODÜLLERİNİN SIVI BAZLI (SurePath) SİTOLOJİSİ: 3 Yıllık Histopatolojik Korelasyon Dr. M. Salih Deveci GATA Gülhane Askeri Tıp Fakültesi Patoloji AD, Ankara Patoloji & Sitopatoloji Kongresi Bursa
TİROİD NODÜLLERİNİN SIVI BAZLI (SurePath) SİTOLOJİSİ: 3 Yıllık Histopatolojik Korelasyon Dr. M. Salih Deveci GATA Gülhane Askeri Tıp Fakültesi Patoloji AD, Ankara Patoloji & Sitopatoloji Kongresi Bursa
Teori (saat/hafta) Laboratuar (saat/hafta) BES114 2. BAHAR 3 0 0 2
 Laboratuar (saat/hafta) BES114 2. BAHAR 3 0 0 2") TIBBİ BİYOLOJİ VE GENETİK Dersin Adı Kodu Yarıyıl TIBBİ BİYOLOJİ VE GENETİK Önkoşullar Dersin dili Dersin Türü Dersin öğrenme ve öğretme teknikleri Dersin sorumlusu(ları) Dersin amacı Dersin öğrenme çıktıları
TIBBİ BİYOLOJİ VE GENETİK Dersin Adı Kodu Yarıyıl TIBBİ BİYOLOJİ VE GENETİK Önkoşullar Dersin dili Dersin Türü Dersin öğrenme ve öğretme teknikleri Dersin sorumlusu(ları) Dersin amacı Dersin öğrenme çıktıları
I. YARIYIL MOLEKÜLER HÜCRE BİYOLOJİSİ I (TBG 601, ZORUNLU, TEORİK 3, 3 KREDİ)
") T. C. İSTANBUL BİLİM ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ VE GENETİK ANABİLİM DALI YÜKSEK LİSANS PROGRAMI 2017-2018 EĞİTİM-ÖĞRETİM YILI DERS İÇERİKLERİ I. YARIYIL MOLEKÜLER HÜCRE BİYOLOJİSİ
T. C. İSTANBUL BİLİM ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TIBBİ BİYOLOJİ VE GENETİK ANABİLİM DALI YÜKSEK LİSANS PROGRAMI 2017-2018 EĞİTİM-ÖĞRETİM YILI DERS İÇERİKLERİ I. YARIYIL MOLEKÜLER HÜCRE BİYOLOJİSİ
MİDE KANSERİNDE P53 EKSPRESYONUNUN PROGNOSTİK ÖNEMİ: META- ANALİZ
 MİDE KANSERİNDE P53 EKSPRESYONUNUN PROGNOSTİK ÖNEMİ: META- ANALİZ Mustafa Yıldırım 1, Vildan Kaya 2, Özlem Demirpençe 3, Hakan Bozcuk 4 1-T.C. Sağlık Bakanlığı Türkiye Kamu Hastaneleri Kurumu Batman İli
MİDE KANSERİNDE P53 EKSPRESYONUNUN PROGNOSTİK ÖNEMİ: META- ANALİZ Mustafa Yıldırım 1, Vildan Kaya 2, Özlem Demirpençe 3, Hakan Bozcuk 4 1-T.C. Sağlık Bakanlığı Türkiye Kamu Hastaneleri Kurumu Batman İli
ÇANAKKALE ONSEKİZ MART ÜNİVERSİTESİ TIP FAKÜLTESİ
 Dönem V Tıbbi Genetik Staj Eğitim Programı Eğitim Başkoordinatörü: Dönem Koordinatörü: Koordinatör Yardımcısı: Doç. Dr. Erkan Melih ŞAHİN Baran GENCER Oğuz GÜÇLÜ Erkam KÖMÜRCÜ Staj Eğitim Sorumlusu: Genel
Dönem V Tıbbi Genetik Staj Eğitim Programı Eğitim Başkoordinatörü: Dönem Koordinatörü: Koordinatör Yardımcısı: Doç. Dr. Erkan Melih ŞAHİN Baran GENCER Oğuz GÜÇLÜ Erkam KÖMÜRCÜ Staj Eğitim Sorumlusu: Genel